
- Current
- Browse
- Collections
-
For contributors
- For Authors
- Instructions to authors
- Article processing charge
- e-submission
- For Reviewers
- Instructions for reviewers
- How to become a reviewer
- Best reviewers
- For Readers
- Readership
- Subscription
- Permission guidelines
- About
- Editorial policy
Articles
- Page Path
- HOME > Diabetes Metab J > Volume 36(3); 2012 > Article
-
ReviewOthers Retinoid Metabolism and Diabetes Mellitus
- Eun-Jung Rhee1, Jorge Plutzky2
-
Diabetes & Metabolism Journal 2012;36(3):167-180.
DOI: https://doi.org/10.4093/dmj.2012.36.3.167
Published online: June 14, 2012
1Department of Endocrinology and Metabolism, Kangbuk Samsung Hospital, Sungkyunkwan University School of Medicine, Seoul, Korea.
2Cardiovascular Division, Brigham and Women's Hospital, Harvard Medical School, Boston, MA, USA.
- Corresponding author: Jorge Plutzky. Cardiovascular Division, Brigham and Women's Hospital, Harvard Medical School, 77 Avenue Louis Pasteur, NRB 742, Boston, MA 02115, USA. jplutzky@rics.bwh.harvard.edu
Copyright © 2012 Korean Diabetes Association
This is an Open Access article distributed under the terms of the Creative Commons Attribution Non-Commercial License (http://creativecommons.org/licenses/by-nc/3.0/) which permits unrestricted non-commercial use, distribution, and reproduction in any medium, provided the original work is properly cited.
- ABSTRACT
- INTRODUCTION
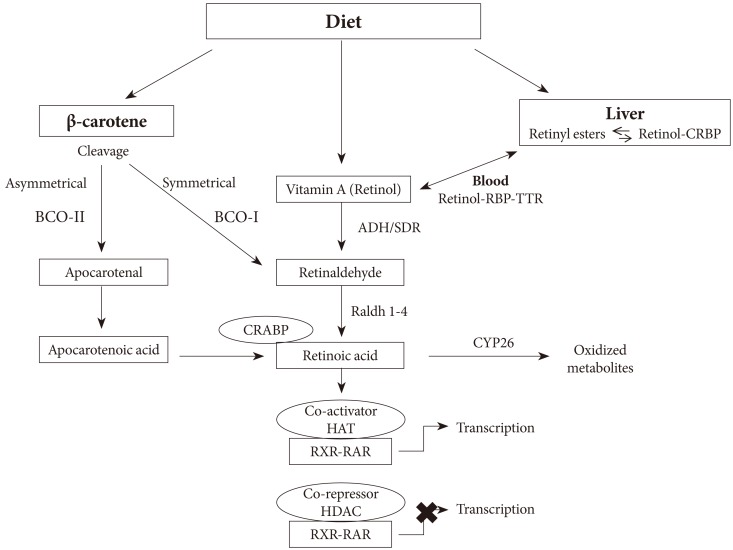
- OVERVIEW OF RETINOIC ACID METABOLISM
- RETINOID METABOLISM AND DIABETES-EVIDENCE FROM HUMAN STUDIES
- RXR-PPAR-γ HETERODIMER: TARGETS IN METABOLIC DISEASES
- RETINOID METABOLISM AND INSULIN SECRETION
- EFFECTS OF RETINOID ON THE PREVENTION OF TYPE 1 DIABETES THROUGH THE MODULATION OF IMMUNE FUNCTION
- RETINOID METABOLISM AND INSULIN SENSITIVITY
- RETINOID METABOLISM AND HEPATIC GLUCONEOGENESIS
- RBP4 AND DIABETES
- THE EFFECTS OF RETINOID THERAPY ON INSULIN SENSITIVITY AND LIPID PROFILES IN HUMANS
- RETINOID METABOLISM AND LIPID METABOLISM
- CONCLUSIONS
- NOTES
- REFERENCES
- About this article
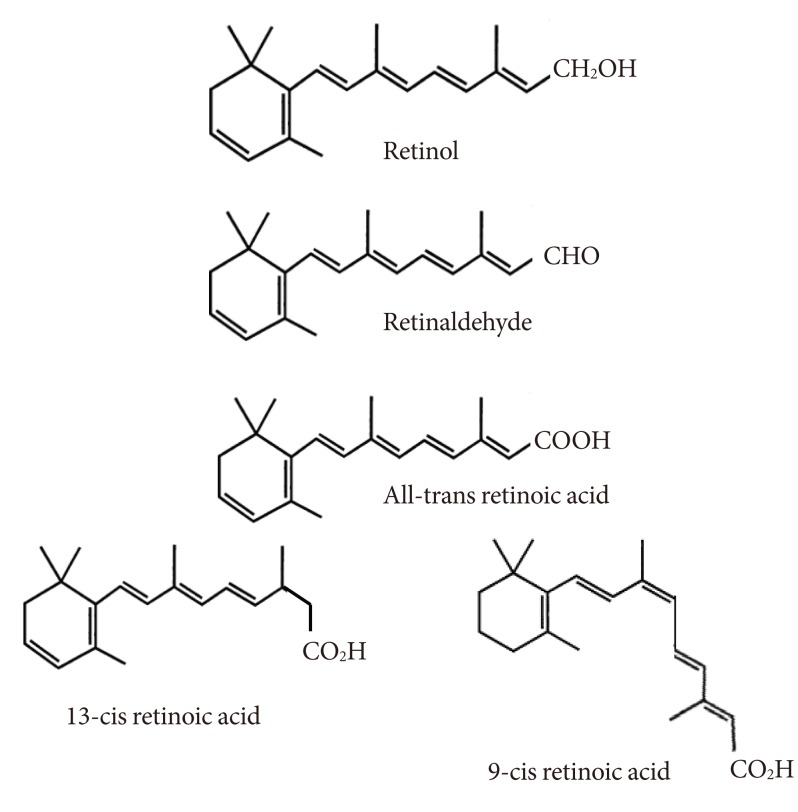
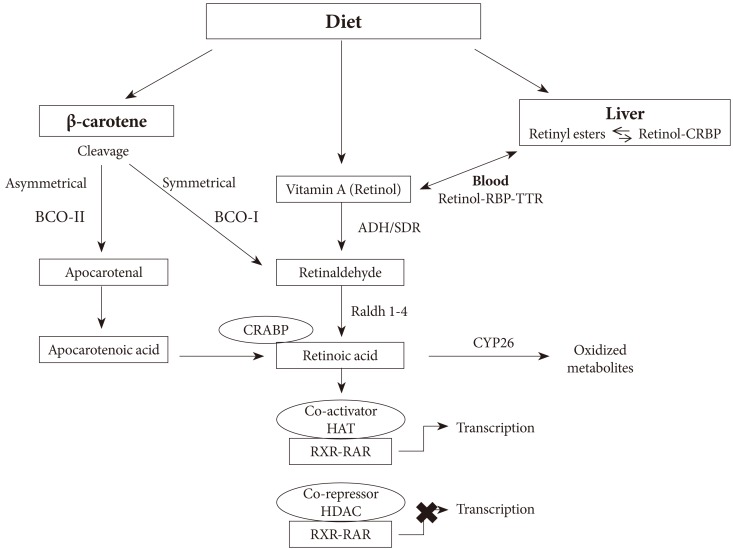
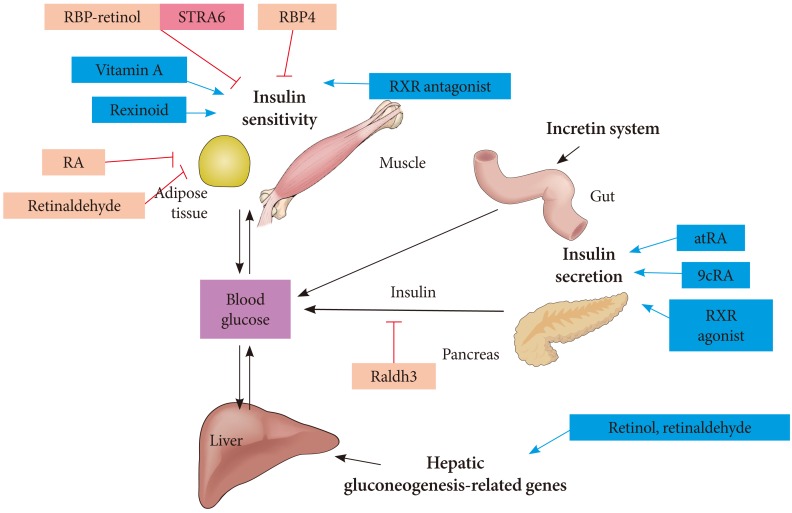
Figure & Data
References
Citations
Citations to this article as recorded by 
- Dysregulated cysteine metabolism leads to worsened liver pathology in diabetes-tuberculosis comorbid condition
Shweta Chaudhary, Falak Pahwa, Ranjan K. Nanda
Journal of Biological Chemistry.2024; 300(2): 105634. CrossRef - Role of dietary antioxidants in diabetes: An overview
Mohamed Shafras, Rasangi Sabaragamuwa, Mohamed Suwair
Food Chemistry Advances.2024; 4: 100666. CrossRef - Characterization of the gut bacterial and viral microbiota in latent autoimmune diabetes in adults
Casper S. Poulsen, Dan Hesse, Gabriel R. Fernandes, Tue H. Hansen, Timo Kern, Allan Linneberg, Lore Van Espen, Torben Jørgensen, Trine Nielsen, Amra C. Alibegovic, Jelle Matthijnssens, Oluf Pedersen, Henrik Vestergaard, Torben Hansen, Mette K. Andersen
Scientific Reports.2024;[Epub] CrossRef - Positive impacts of Nannochloropsis oculata supplementation on gene expression of immune and antioxidant markers and metabolic profile of Barki sheep in the transition period and lipogenic effects on progeny
Ahmed El-Sayed, Eman Ebissy, Ahmed Ateya
Veterinary Research Communications.2024;[Epub] CrossRef - Association of plasma vitamin A level with type 2 diabetes mellitus: a community aging population-based cross-sectional study
Pengfei Li, Jingjing Xu, Yujie Guo, Xiaojun Ma, Shaobo Zhou, Chi Zhang, Huiyan Yu, Ying Wang, Xixiang Wang, Linhong Yuan
Food Science and Human Wellness.2024; 13(4): 2079. CrossRef - Retinoic acid signaling is critical for generation of pancreatic progenitors from human embryonic stem cells
Niloufer P. Dumasia, Aparna P. Khanna, Prasad S. Pethe
Growth Factors.2023; 41(1): 8. CrossRef - Immuno-metabolic control of the balance between Th17-polarized and regulatory T-cells during HIV infection
Alexis Yero, Ralph-Sydney Mboumba Bouassa, Petronela Ancuta, Jerome Estaquier, Mohammad-Ali Jenabian
Cytokine & Growth Factor Reviews.2023; 69: 1. CrossRef - The transcriptomic and epigenetic alterations in type 2 diabetes mellitus patients of Chinese Tibetan and Han populations
Xian Wang, Jie Liu, Qiuhong Wang, Qiu Chen
Frontiers in Endocrinology.2023;[Epub] CrossRef - Pericytes modulate islet immune cells and insulin secretion through Interleukin-33 production in mice
Guzel Burganova, Anat Schonblum, Lina Sakhneny, Alona Epshtein, Tomer Wald, Mika Tzaig, Limor Landsman
Frontiers in Endocrinology.2023;[Epub] CrossRef - Marine-Derived Compounds Applied in Cardiovascular Diseases: Submerged Medicinal Industry
Wasim Akram, Mohd Rihan, Sakeel Ahmed, Swamita Arora, Sameer Ahmad, Rahul Vashishth
Marine Drugs.2023; 21(3): 193. CrossRef - Metabolite Profiling of the Gut–Renal–Cerebral Axis Reveals a Particular Pattern in Early Diabetic Kidney Disease in T2DM Patients
Lavinia Balint, Carmen Socaciu, Andreea Iulia Socaciu, Adrian Vlad, Florica Gadalean, Flaviu Bob, Oana Milas, Octavian Marius Cretu, Anca Suteanu-Simulescu, Mihaela Glavan, Silvia Ienciu, Maria Mogos, Dragos Catalin Jianu, Ligia Petrica
International Journal of Molecular Sciences.2023; 24(7): 6212. CrossRef - Multimodal plasma metabolomics and lipidomics in elucidating metabolic perturbations in tuberculosis patients with concurrent type 2 diabetes
Nguyen Thi Hai Yen, Nguyen Ky Anh, Rannissa Puspita Jayanti, Nguyen Ky Phat, Dinh Hoa Vu, Jong-Lyul Ghim, Sangzin Ahn, Jae-Gook Shin, Jee Youn Oh, Nguyen Phuoc Long, Dong Hyun Kim
Biochimie.2023; 211: 153. CrossRef - Microbiota dysbiosis associated with type 2 diabetes-like effects caused by chronic exposure to a mixture of chlorinated persistent organic pollutants in zebrafish
Hyojin Lee, Sojeong Yoon, Youngja Hwang Park, Jeong-Soo Lee, Dong Young Rhyu, Ki-Tae Kim
Environmental Pollution.2023; 334: 122108. CrossRef - Hepatic retinaldehyde deficiency is involved in diabetes deterioration by enhancing PCK1- and G6PC-mediated gluconeogenesis
Hanyu Yang, Mengxiang Su, Ming Liu, Yun Sheng, Liang Zhu, Lu Yang, Ruijing Mu, Jianjun Zou, Xiaodong Liu, Li Liu
Acta Pharmaceutica Sinica B.2023; 13(9): 3728. CrossRef - The Association and Mediating Biomarkers of Serum Retinol in Influencing the Development of Type 2 Diabetes: A Prospective Cohort Study in Middle-Aged and Elderly Population
Xiuyu Pang, Sen Yang, Xiaoyu Guo, Hongyin Li, Yingfeng Zhang, Chunbo Wei, Yu Wang, Changhao Sun, Ying Li
Frontiers in Nutrition.2022;[Epub] CrossRef - Quantification of All-Trans Retinoic Acid by Liquid Chromatography–Tandem Mass Spectrometry and Association with Lipid Profile in Patients with Type 2 Diabetes
Jakob Morgenstern, Thomas Fleming, Elisabeth Kliemank, Maik Brune, Peter Nawroth, Andreas Fischer
Metabolites.2021; 11(1): 60. CrossRef - Vitamin A and Its Multi-Effects on Pancreas: Recent Advances and Prospects
Yunting Zhou, Huiying Wang, Junming Zhou, Shanhu Qiu, Tingting Cai, Huiqin Li, Ziyang Shen, Yun Hu, Bo Ding, Menghui Luo, Rong Huang, Rengna Yan, Wei Xu, Cong He, Yumin Zhang, Fengfei Li, Zilin Sun, Jianhua Ma
Frontiers in Endocrinology.2021;[Epub] CrossRef - Association between Vitamin A, E and Type 2 Diabetes Mellitus in Korea: Korean National Health and Nutrition Examination Survey 2016–2018
Kyungpyo Kim, Junho Choi, Serng Bai Pak
Korean Journal of Family Practice.2021; 11(2): 135. CrossRef - Serum Retinal and Retinoic Acid Predict the Development of Type 2 Diabetes Mellitus in Korean Subjects with Impaired Fasting Glucose from the KCPS-II Cohort
Youngmin Han, Yeunsoo Yang, Minjoo Kim, Sun Ha Jee, Hye Jin Yoo, Jong Ho Lee
Metabolites.2021; 11(8): 510. CrossRef - The Interactions of Insulin and Vitamin A Signaling Systems for the Regulation of Hepatic Glucose and Lipid Metabolism
Guoxun Chen
Cells.2021; 10(8): 2160. CrossRef - RETSAT Mutation Selected for Hypoxia Adaptation Inhibits Tumor Growth
Xiulin Jiang, Yaomei He, Qiushuo Shen, Lincan Duan, Yixiao Yuan, Lin Tang, Yulin Shi, Baiyang Liu, Haoqing Zhai, Peng Shi, Cuiping Yang, Yongbin Chen
Frontiers in Cell and Developmental Biology.2021;[Epub] CrossRef - Research Progress in Vitamin A and Autism Spectrum Disorder
Zhonghui Liu, Jingyu Wang, Qu Xu, Qin Hong, Jiansheng Zhu, Xia Chi, Enzo Emanuele
Behavioural Neurology.2021; 2021: 1. CrossRef - Reductive Effect of Acitretin on Blood Glucose Levels in Chinese Patients With Psoriasis
Hua Qian, Yehong Kuang, Juan Su, Menglin Chen, Xiang Chen, Chengzhi Lv, Wangqing Chen, Wu Zhu
Frontiers in Medicine.2021;[Epub] CrossRef - Preventive action of retinoids in metabolic syndrome/type 2 diabetic rats fed with citrus functional food enriched in β-cryptoxanthin
Claudie Dhuique-Mayer, Laura Gence, Karine Portet, Didier Tousch, Patrick Poucheret
Food & Function.2020; 11(10): 9263. CrossRef - CNV Association of Diverse Clinical Phenotypes from eMERGE reveals novel disease biology underlying cardiovascular disease
Joseph T. Glessner, Jin Li, Akshatha Desai, Melody Palmer, Dokyoon Kim, Anastasia Marie Lucas, Xiao Chang, John J. Connolly, Berta Almoguera, John B. Harley, Gail P. Jarvik, Marylyn D. Ritchie, Patrick M.A. Sleiman, Dan M. Roden, David Crosslin, Hakon Hak
International Journal of Cardiology.2020; 298: 107. CrossRef - Pancreatic stellate cell‐potentiated insulin secretion from Min6 cells is independent of interleukin 6‐mediated pathway
Ratnakar R. Bynigeri, Sasikala Mitnala, Rupjyoti Talukdar, Surya S. Singh, Nageshwar R. Duvvuru
Journal of Cellular Biochemistry.2020; 121(1): 840. CrossRef - Impaired CPT1A Gene Expression Response to Retinoic Acid Treatment in Human PBMC as Predictor of Metabolic Risk
Margalida Cifre, Andreu Palou, Paula Oliver
Nutrients.2020; 12(8): 2269. CrossRef - All-Trans Retinoic Acid Enhances both the Signaling for Priming and the Glycolysis for Activation of NLRP3 Inflammasome in Human Macrophage
Ahmad Alatshan, Gergő E. Kovács, Azzam Aladdin, Zsolt Czimmerer, Krisztina Tar, Szilvia Benkő
Cells.2020; 9(7): 1591. CrossRef - Genetic immune and inflammatory markers associated with diabetes in solid organ transplant recipients
Lina Quteineh, Agnieszka Wójtowicz, Pierre-Yves Bochud, Severine Crettol, Frederik Vandenberghe, Jean-Pierre Venetz, Oriol Manuel, Dela Golshayan, Roger Lehmann, Nicolas J. Mueller, Isabelle Binet, Christian van Delden, Jürg Steiger, Paul Mohacsi, Jean-Fr
American Journal of Transplantation.2019; 19(1): 238. CrossRef - Vitamin A and Wound Healing
Roman Zinder, Rachel Cooley, Lucian G. Vlad, Joseph A. Molnar
Nutrition in Clinical Practice.2019; 34(6): 839. CrossRef - In utero exposure to bisphenol-A disrupts key elements of retinoid system in male mice offspring
Javier Esteban, Marina Serrano-Maciá, Ismael Sánchez-Pérez, Paloma Alonso-Magdalena, María de la Cruz Pellín, Marta García-Arévalo, Ángel Nadal, Jose Barril
Food and Chemical Toxicology.2019; 126: 142. CrossRef -
Risk variants disrupting enhancers of T
H
1 and T
REG
cells in type 1 diabetes
Peng Gao, Yasin Uzun, Bing He, Sarah E. Salamati, Julie K. M. Coffey, Eva Tsalikian, Kai Tan
Proceedings of the National Academy of Sciences.2019; 116(15): 7581. CrossRef - Astaxanthin: A Potential Mitochondrial-Targeted Antioxidant Treatment in Diseases and with Aging
Mónika Sztretye, Beatrix Dienes, Mónika Gönczi, Tamás Czirják, László Csernoch, László Dux, Péter Szentesi, Anikó Keller-Pintér
Oxidative Medicine and Cellular Longevity.2019; 2019: 1. CrossRef - Design and rationale of the INSYTE study: A randomised, placebo controlled study to test the efficacy of a synbiotic on liver fat, disease biomarkers and intestinal microbiota in non-alcoholic fatty liver disease
Eleonora Scorletti, Paul R. Afolabi, Elizabeth A. Miles, Debbie E. Smith, Amal Almehmadi, Albandri Alshathry, Helen E. Moyses, Geraldine F. Clough, Mark Wright, Janisha Patel, Laure Bindels, Nathalie M. Delzenne, Philip C. Calder, Christopher D. Byrne
Contemporary Clinical Trials.2018; 71: 113. CrossRef - Protective effect of supplementation with biotin against high-fructose-induced metabolic syndrome in rats
Asdrubal Aguilera-Mendez, María G. Hernández-Equihua, Alfonso C. Rueda-Rocha, Clotilde Guajardo-López, Renato Nieto-Aguilar, Deyanira Serrato-Ochoa, León F. Ruíz Herrera, José A. Guzmán-Nateras
Nutrition Research.2018; 57: 86. CrossRef - Elevated Fibroblast growth factor 21 (FGF21) in obese, insulin resistant states is normalised by the synthetic retinoid Fenretinide in mice
Nicola Morrice, George D. Mcilroy, Seshu R. Tammireddy, Jennifer Reekie, Kirsty D. Shearer, Mary K. Doherty, Mirela Delibegović, Phillip D. Whitfield, Nimesh Mody
Scientific Reports.2017;[Epub] CrossRef - Vitamin supplements in type 2 diabetes mellitus management: A review
Mabel Kar-Wai Yan, Hanan Khalil
Diabetes & Metabolic Syndrome: Clinical Research & Reviews.2017; 11: S589. CrossRef - Isotretinoin als „second hit“ bei Schizophrenie?
Michael Kaczmarczyk, Carolin Laqua, Julian Hellmann-Regen, Isabella Heuser, Francesca Regen, Eric Hahn
NeuroTransmitter.2017; 28(1): 29. CrossRef - Role of vitamin A metabolism in IIH: Results from the idiopathic intracranial hypertension treatment trial
J Libien, MJ Kupersmith, W Blaner, MP McDermott, S Gao, Y Liu, J Corbett, M Wall
Journal of the Neurological Sciences.2017; 372: 78. CrossRef - Synthesis of a new insulin-mimetic anti-diabetic drug containing vitamin A and vanadium(IV) salt: Chemico-biological characterizations
Abdel Majid A Adam, Ahmed M Naglah, Mohamed A Al-Omar, Moamen S Refat
International Journal of Immunopathology and Pharmacology.2017; 30(3): 272. CrossRef - The LC–MS-based metabolomics of hydroxytyrosol administration in rats reveals amelioration of the metabolic syndrome
Nikolaos Lemonakis, Hemant Poudyal, Maria Halabalaki, Lindsay Brown, Anthony Tsarbopoulos, Alexios-Leandros Skaltsounis, Evagelos Gikas
Journal of Chromatography B.2017; 1041-1042: 45. CrossRef - High serum carotenoids are associated with lower risk for developing elevated serum alanine aminotransferase among Japanese subjects: the Mikkabi cohort study
Minoru Sugiura, Mieko Nakamura, Kazunori Ogawa, Yoshinori Ikoma, Masamichi Yano
British Journal of Nutrition.2016; 115(8): 1462. CrossRef - Retinoids have therapeutic action in type 2 diabetes
Dilnasheen Meerza, Sarah Iqbal, Sufian Zaheer, Imrana Naseem
Nutrition.2016; 32(7-8): 898. CrossRef - Altered retinol status and expression of retinol-related proteins in streptozotocin-induced type 1 diabetic model rats
Kimitaka Takitani, Keisuke Inoue, Maki Koh, Hiroshi Miyazaki, Akiko Inoue, Kanta Kishi, Hiroshi Tamai
Journal of Clinical Biochemistry and Nutrition.2015; 56(3): 195. CrossRef - Role of vitamin A in type 2 diabetes mellitus biology: Effects of intervention therapy in a deficient state
Sarah Iqbal, Imrana Naseem
Nutrition.2015; 31(7-8): 901. CrossRef - Prepartum vitamin A supplementation enhances goat doe health status and kid viability and performance
Mabrouk A. Abd Eldaim, Khalid M. Gaafar, Ragab A. Darwish, Hamad D. Mahboub, Mohamed A. Helal
Small Ruminant Research.2015; 129: 6. CrossRef - Beta Cell Function and the Nutritional State: Dietary Factors that Influence Insulin Secretion
William T. Moore, Suzanne M. Bowser, Dane W. Fausnacht, Linda L. Staley, Kyung-Shin Suh, Dongmin Liu
Current Diabetes Reports.2015;[Epub] CrossRef - High-serum carotenoids associated with lower risk for developing type 2 diabetes among Japanese subjects: Mikkabi cohort study
Minoru Sugiura, Mieko Nakamura, Kazunori Ogawa, Yoshinori Ikoma, Masamichi Yano
BMJ Open Diabetes Research & Care.2015; 3(1): e000147. CrossRef - Gestational diabetes: a risk of puerperal hypovitaminosis A?
FERNANDA B.S. RESENDE, LARISSA Q. DE LIRA, EVELLYN C. GRILO, MAYARA S.R. LIMA, ROBERTO DIMENSTEIN
Anais da Academia Brasileira de Ciências.2015; 87(1): 463. CrossRef - Plasma concentrations of retinol in obese children and adolescents: relationship to metabolic syndrome components
Marcia Teske, Ana Paula B. Melges, Fabiola Isabel S. de Souza, Fernando Luiz A. Fonseca, Roseli Oselka S. Sarni
Revista Paulista de Pediatria.2014; 32(1): 50. CrossRef - Proteomic analysis of livers from fat-fed mice deficient in either PKCδ or PKCε identifies Htatip2 as a regulator of lipid metabolism
Bing M. Liao, Katy Raddatz, Ling Zhong, Benjamin L. Parker, Mark J. Raftery, Carsten Schmitz-Peiffer
PROTEOMICS.2014; 14(21-22): 2578. CrossRef - Effect of all-trans-retinoic acid on the structure of thyroid gland and pituitary tyrotrophs in streptozotocin-diabetic male rats
Ola A. Hussein, Amel M.M. Abdel-Hafez
The Egyptian Journal of Histology.2013; 36(3): 691. CrossRef - Quantitative Proteomic and Functional Analysis of Liver Mitochondria from High Fat Diet (HFD) Diabetic Mice
Yurong Guo, Manjula Darshi, Yuliang Ma, Guy A. Perkins, Zhouxin Shen, Kristofer J. Haushalter, Rintaro Saito, Ai Chen, Yun Sok Lee, Hemal H. Patel, Steven P. Briggs, Mark H. Ellisman, Jerrold M. Olefsky, Susan S. Taylor
Molecular & Cellular Proteomics.2013; 12(12): 3744. CrossRef - Deriving at candidate genes of metabolic stress from pancreas of WNIN/GR-Ob mutant rats
Himadri Singh, Nappanveettil Giridharan, Ramesh Bhonde, Vijayalakshmi Venkatesan
Islets.2013; 5(4): 133. CrossRef
 PubReader
PubReader Cite this Article
Cite this Article
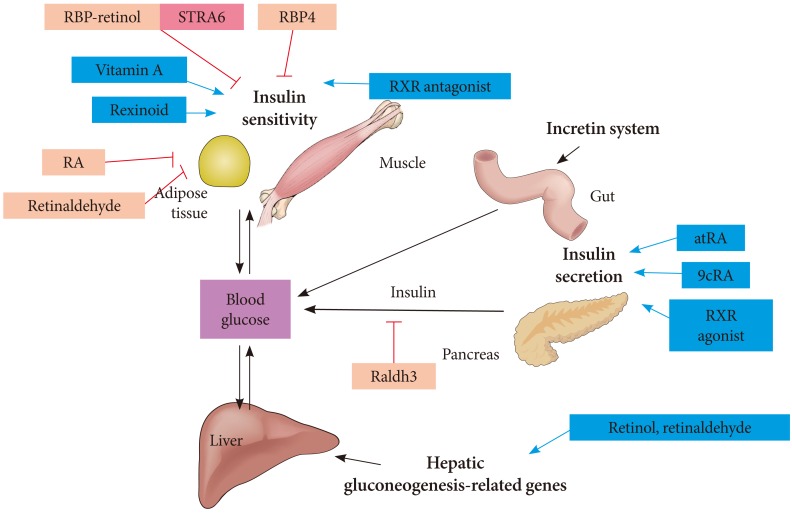
- Rhee EJ, Plutzky J. Retinoid Metabolism and Diabetes Mellitus. Diabetes Metab J. 2012;36(3):167-180.
- DOI: https://doi.org/10.4093/dmj.2012.36.3.167.
-
- Download citation ↓




