- Current
- Browse
- Collections
-
For contributors
- For Authors
- Instructions to authors
- Article processing charge
- e-submission
- For Reviewers
- Instructions for reviewers
- How to become a reviewer
- Best reviewers
- For Readers
- Readership
- Subscription
- Permission guidelines
- About
- Editorial policy
Articles
- Page Path
- HOME > Diabetes Metab J > Volume 34(5); 2010 > Article
-
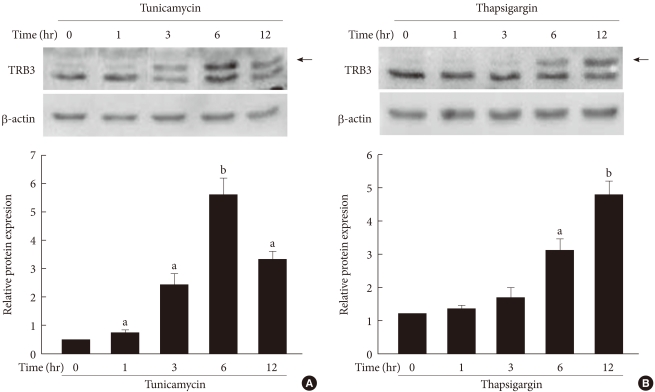
Original ArticleThe Effect of Tribbles-Related Protein 3 on ER Stress-Suppressed Insulin Gene Expression in INS-1 Cells
- Young Yun Jang1, Nam Keong Kim1, Mi Kyung Kim1, Ho Young Lee1, Sang Jin Kim1, Hye Soon Kim1, Hye-Young Seo2, In Kyu Lee2, Keun Gyu Park1
-
Korean Diabetes Journal 2010;34(5):312-319.
DOI: https://doi.org/10.4093/kdj.2010.34.5.312
Published online: October 31, 2010
- 3,592 Views
- 32 Download
- 5 Crossref
1Department of Internal Medicine, Keimyung University School of Medicine, Daegu, Korea.
2Department of Internal Medicine, Kyungpook National University School of Medicine, Daegu, Korea.
- Corresponding author: Keun Gyu Park. Department of Internal Medicine, Keimyung University School of Medicine, 194 Dongsan-dong, Jung-gu, Daegu 700-712, Korea. kgpark@dsmc.or.kr
• Received: March 23, 2010 • Accepted: August 11, 2010
Copyright © 2010 Korean Diabetes Association
This is an Open Access article distributed under the terms of the Creative Commons Attribution Non-Commercial License (http://creativecommons.org/licenses/by-nc/3.0/) which permits unrestricted non-commercial use, distribution, and reproduction in any medium, provided the original work is properly cited.
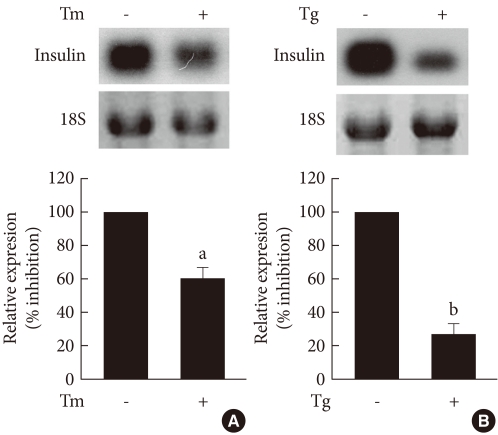
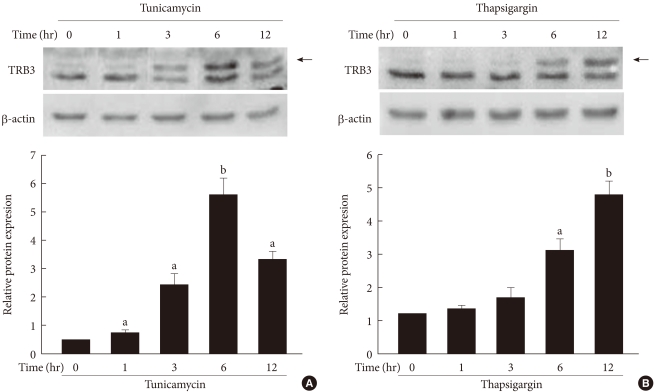
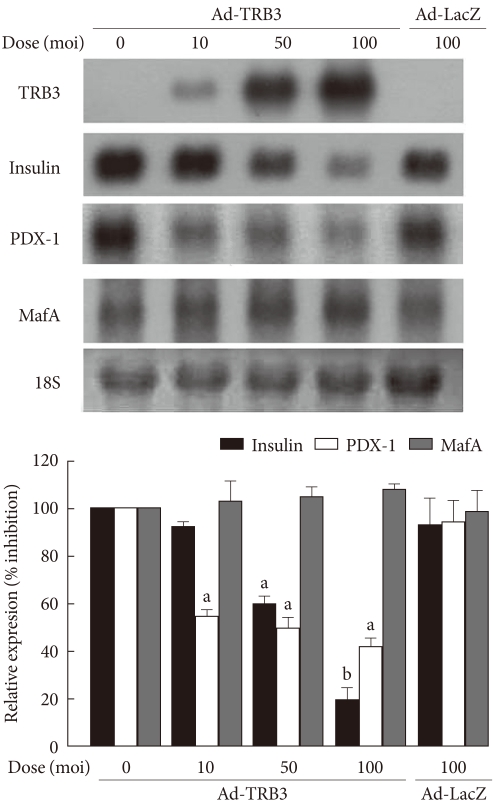
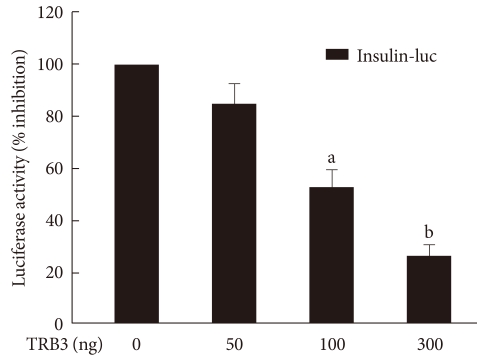
Figure & Data
References
Citations
Citations to this article as recorded by 
- Endoplasmic reticulum stress causes insulin resistance by inhibiting delivery of newly synthesized insulin receptors to the cell surface
Max Brown, Samantha Dainty, Natalie Strudwick, Adina D. Mihai, Jamie N. Watson, Robina Dendooven, Adrienne W. Paton, James C. Paton, Martin Schröder, James Arthur Olzmann
Molecular Biology of the Cell.2020; 31(23): 2597. CrossRef - PTB and TIAR binding to insulin mRNA 3′- and 5′UTRs; implications for insulin biosynthesis and messenger stability
Rikard G. Fred, Syrina Mehrabi, Christopher M. Adams, Nils Welsh
Heliyon.2016; 2(9): e00159. CrossRef - Asna1/TRC40 Controls β-Cell Function and Endoplasmic Reticulum Homeostasis by Ensuring Retrograde Transport
Stefan Norlin, Vishal S. Parekh, Peter Naredi, Helena Edlund
Diabetes.2016; 65(1): 110. CrossRef - Role of the Unfolded Protein Response inβCell Compensation and Failure during Diabetes
Nabil Rabhi, Elisabet Salas, Philippe Froguel, Jean-Sébastien Annicotte
Journal of Diabetes Research.2014; 2014: 1. CrossRef - Endoplasmic Reticulum Stress and Insulin Biosynthesis: A Review
Mi-Kyung Kim, Hye-Soon Kim, In-Kyu Lee, Keun-Gyu Park
Experimental Diabetes Research.2012; 2012: 1. CrossRef
 PubReader
PubReader Cite
Cite