- Current
- Browse
- Collections
-
For contributors
- For Authors
- Instructions to authors
- Article processing charge
- e-submission
- For Reviewers
- Instructions for reviewers
- How to become a reviewer
- Best reviewers
- For Readers
- Readership
- Subscription
- Permission guidelines
- About
- Editorial policy
Articles
- Page Path
- HOME > Diabetes Metab J > Volume 44(2); 2020 > Article
-
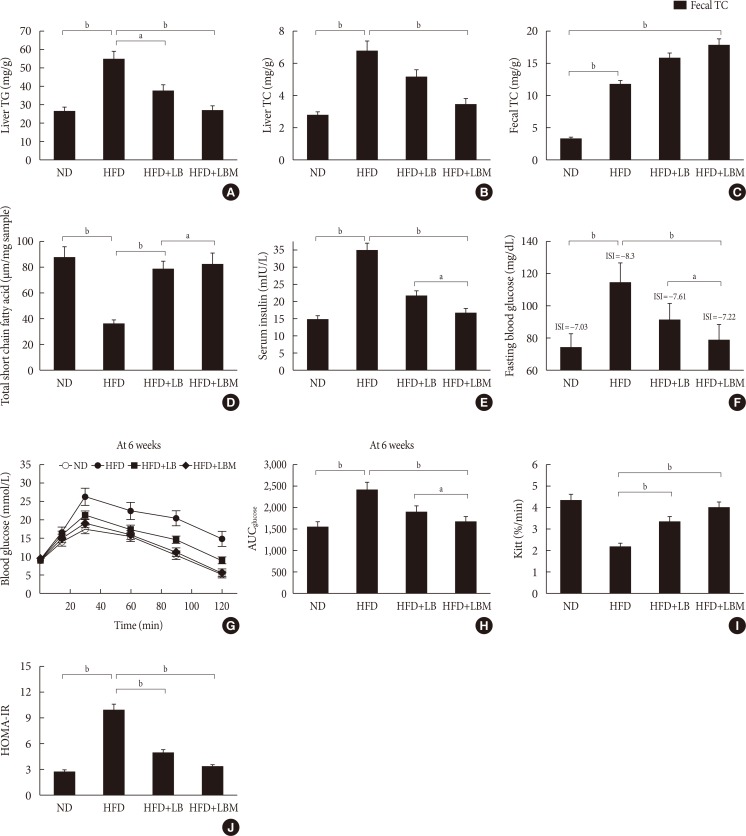
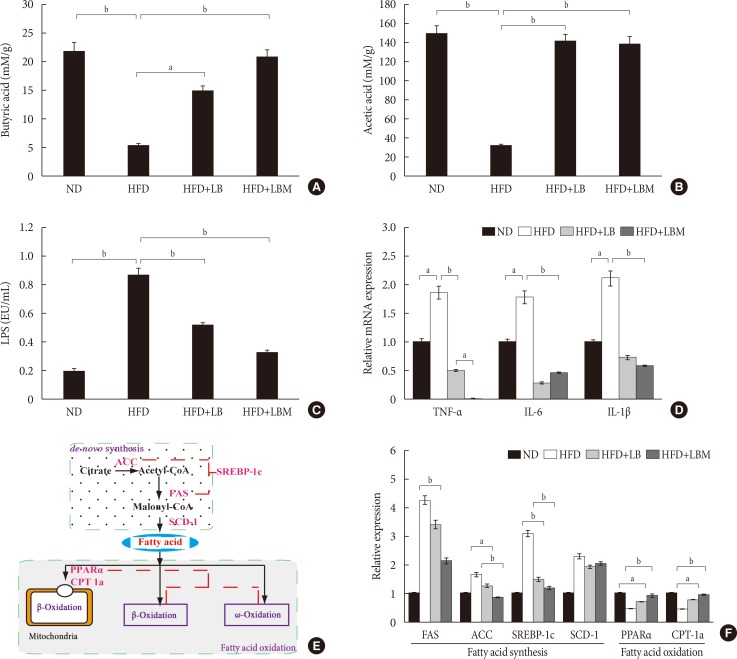
Original ArticleBasic Research Combination of Probiotics and
Salvia miltiorrhiza Polysaccharide Alleviates Hepatic Steatosis via Gut Microbiota Modulation and Insulin Resistance Improvement in High Fat-Induced NAFLD Mice -
Wei Wang1,2
 , Ai-Lei Xu2, Zheng-Chao Li1, Yi Li2, Shun-Fu Xu2, Hua-Chao Sang2, Fachao Zhi1
, Ai-Lei Xu2, Zheng-Chao Li1, Yi Li2, Shun-Fu Xu2, Hua-Chao Sang2, Fachao Zhi1 -
Diabetes & Metabolism Journal 2020;44(2):336-348.
DOI: https://doi.org/10.4093/dmj.2019.0042
Published online: December 3, 2019
1Guangdong Provincial Key Laboratory of Gastroenterology, Department of Gastroenterology, Institute of Gastroenterology of Guangdong Province, Nanfang Hospital, Southern Medical University, Guangzhou, China.
2Department of Gastroenterology, Sir Run Run Hospital, Nanjing Medical University, Nanjing, China.
- Corresponding author: Fachao Zhi. Guangdong Provincial Key Laboratory of Gastroenterology, Department of Gastroenterology, Institute of Gastroenterology of Guangdong Province, Nanfang Hospital, Southern Medical University, Guangzhou, No. 1838, Guangzhou Avenue North Rd, Baiyun District, Guangzhou 510515, China. zhifc41532@163.com
• Received: March 4, 2019 • Accepted: May 9, 2019
Copyright © 2020 Korean Diabetes Association
This is an Open Access article distributed under the terms of the Creative Commons Attribution Non-Commercial License (http://creativecommons.org/licenses/by-nc/4.0/) which permits unrestricted non-commercial use, distribution, and reproduction in any medium, provided the original work is properly cited.

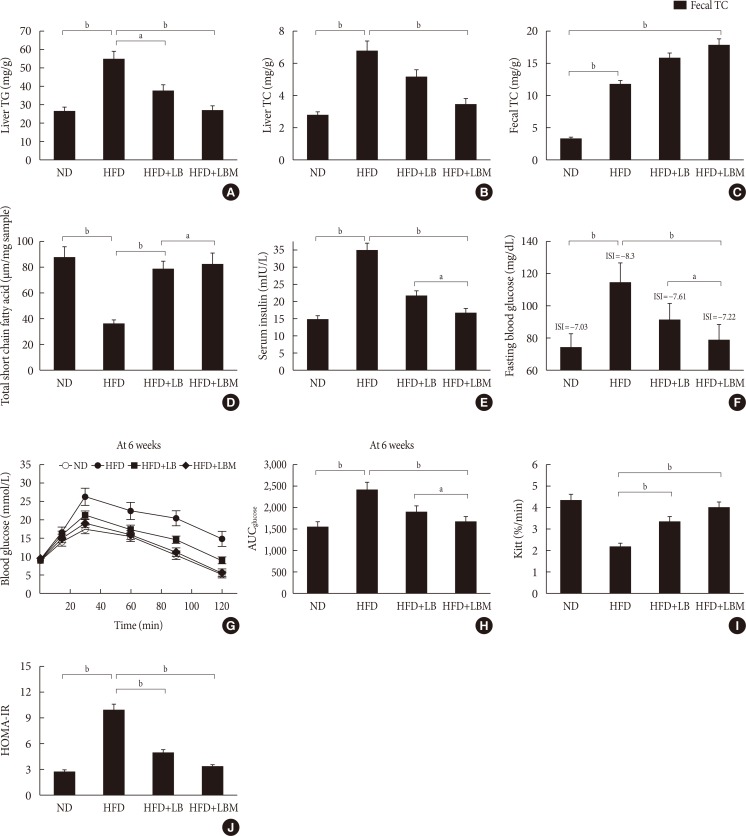


Figure & Data
References
Citations
Citations to this article as recorded by 
- Bifidobacterium bifidum BGN4 fractions ameliorate palmitic acid-induced hepatocyte ferroptosis by inhibiting SREBP1-CYP2E1 pathway
Guangkui Bu, Gang Chen, Juan Li, Dan Wu, Jiangtao Liao
Journal of Investigative Medicine.2024; 72(1): 67. CrossRef - Docosahexaenoic acid (DHA) alleviates hepatic lipid accumulation by regulating mitochondrial quality control through ERK signaling pathway in grass carp (Ctenopharyngodon idellus)
Chenchen Bian, Shanghong Ji, Caihong Zeng, Jian Sun, Gen Kaneko, Hong Ji
Aquaculture.2024; 579: 740209. CrossRef - Microbial-Based Bioactive Compounds to Alleviate Inflammation in Obesity
Oladayo Emmanuel Apalowo, Grace Adeola Adegoye, Tolulope Mobolaji Obuotor
Current Issues in Molecular Biology.2024; 46(3): 1810. CrossRef - Understanding the role of ursodeoxycholic acid and gut microbiome in non-alcoholic fatty liver disease: current evidence and perspectives
Qingyi Mao, Beibei Lin, Wenluo Zhang, Yu Zhang, Yu Zhang, Qian Cao, Mengque Xu
Frontiers in Pharmacology.2024;[Epub] CrossRef - Smilax China L. polysaccharide prevents HFD induced-NAFLD by regulating hepatic fat metabolism and gut microbiota
Wenkai Zhang, Longhui Yu, Qinru Yang, Jinfeng Zhang, Wenjing Wang, Xinru Hu, Jingen Li, Guodong Zheng
Phytomedicine.2024; 127: 155478. CrossRef - Breaking the barriers: the role of gut homeostasis in Metabolic-Associated Steatotic Liver Disease (MASLD)
Raquel Benedé-Ubieto, Francisco Javier Cubero, Yulia A. Nevzorova
Gut Microbes.2024;[Epub] CrossRef - Prospect of research on anti-atherosclerosis effect of main components of traditional Chinese medicine Yiqi Huoxue Huatan recipe through gut microbiota: A review
Hongtao Huang, Hanjun Zhao, Lv Wenqing, Feiyue Xu, Xiaolong Wang, Yili Yao, Yu Huang
Medicine.2024; 103(5): e37104. CrossRef - Polysaccharides from Eucommia ulmoides Oliv. Leaves Alleviate Acute Alcoholic Liver Injury by Modulating the Microbiota–Gut–Liver Axis in Mice
Yingzhi Li, Huimei Wang, Xueping Leng, Jiaming Gao, Chang Li, Danfei Huang
Foods.2024; 13(7): 1089. CrossRef - Innovative pharmacotherapy for hepatic metabolic and chronic inflammatory diseases in China
Feng Zhang, Jiaming Ju, Hongtao Diao, Jinglun Song, Yu bian, Baofeng Yang
British Journal of Pharmacology.2024;[Epub] CrossRef - Shenling Baizhu San ameliorates non-alcoholic fatty liver disease in mice by modulating gut microbiota and metabolites
Dongliang Chen, Yuanfei Wang, Jianmei Yang, Wanyi Ou, Guiru Lin, Ze Zeng, Xiaomin Lu, Zumin Chen, Lili Zou, Yaling Tian, Aiping Wu, Shelley E. Keating, Qinhe Yang, Chenli Lin, Yinji Liang
Frontiers in Pharmacology.2024;[Epub] CrossRef - Intestinal dysbiosis in nonalcoholic fatty liver disease (NAFLD): focusing on the gut–liver axis
Hui Han, Yi Jiang, Mengyu Wang, Mebratu Melaku, Lei Liu, Yong Zhao, Nadia Everaert, Bao Yi, Hongfu Zhang
Critical Reviews in Food Science and Nutrition.2023; 63(12): 1689. CrossRef - Epigenetic compounds targeting pharmacological target lysine specific demethylase 1 and its impact on immunotherapy, chemotherapy and radiotherapy for treatment of tumor recurrence and resistance
Clement Agboyibor, Jianshu Dong, Clement Yaw Effah, Emmanuel Kwateng Drokow, Maxwell Ampomah-Wireko, Waqar Pervaiz, Augustina Sangmor, Xinli Ma, Jian Li, Hong-Min Liu, Peng Zhang
Biomedicine & Pharmacotherapy.2023; 157: 113934. CrossRef - Salvia miltiorrhiza polysaccharides alleviate florfenicol-induced inflammation and oxidative stress in chick livers by regulating phagosome signaling pathway
Xiao Wang, Wei Liu, Di Zhang, Yulan Jiao, Qianhui Zhao, Ying Liu, Wanyu Shi, Yongzhan Bao
Ecotoxicology and Environmental Safety.2023; 249: 114428. CrossRef - Preparation methods, structural characteristics, and biological activity of polysaccharides from Salvia miltiorrhiza: A review
Yuanyuan Li, Xin Zhang, Yining Li, Pei Yang, Zhiyuan Zhang, Hang Wu, Lihao Zhu, Yuhong Liu
Journal of Ethnopharmacology.2023; 305: 116090. CrossRef - The spleen-strengthening and liver-draining herbal formula treatment of non-alcoholic fatty liver disease by regulation of intestinal flora in clinical trial
Dengcheng Hui, Lu Liu, Nisma Lena Bahaji Azami, Jingru Song, Yanping Huang, Wan Xu, Chao Wu, Dong Xie, Yulang Jiang, Yanqin Bian, Mingyu Sun
Frontiers in Endocrinology.2023;[Epub] CrossRef - The gut microbiota: A new perspective for tertiary prevention of hepatobiliary and gallbladder diseases
Xiaoyu Huang, Yi Yang, Xueli Li, Xiaoya Zhu, Dan Lin, Yueran Ma, Min Zhou, Xiangyi Cui, Bingyu Zhang, Dongmei Dang, Yuhong Lü, Changwu Yue
Frontiers in Nutrition.2023;[Epub] CrossRef - Polysaccharides from Ostrea rivularis rebuild the balance of gut microbiota to ameliorate non-alcoholic fatty liver disease in ApoE−/− mice
Lijun Zhu, Mingmei Xiao, Jigang Luo, Shijie Li, Wenting Liu, Jinchuan Wu, Zhuoyue Song
International Journal of Biological Macromolecules.2023; 235: 123853. CrossRef - Improvement of Inflammation, Diabetes, and Obesity by Forest
Product-Derived Polysaccharides through the Human Intestinal
Microbiota
Seong-woo MYEONG, Yong Ju LEE, Do Hyun KIM, Tae-Jong KIM
Journal of the Korean Wood Science and Technology.2023; 51(5): 358. CrossRef - Recent developments in Salvia miltiorrhiza polysaccharides: Isolation, purification, structural characteristics and biological activities
Lei Luo, Juan Xue, Zheng Shao, Zhang Zhou, Wenqian Tang, Jinxin Liu, Hongfei Hu, Fan Yang
Frontiers in Pharmacology.2023;[Epub] CrossRef - In Vitro Probiotic Properties of Bifidobacterium animalis subsp. lactis SF and Its Alleviating Effect on Non-Alcoholic Fatty Liver Disease
Huihui Lv, Feiyue Tao, Lingling Peng, Shufang Chen, Zhongyue Ren, Jiahui Chen, Bo Yu, Hua Wei, Cuixiang Wan
Nutrients.2023; 15(6): 1355. CrossRef - Ginsenoside Rg1 Ameliorates Pancreatic Injuries via the AMPK/mTOR Pathway in vivo and in vitro
Jin Chen, Guoping Zhu, Wenbo Xiao, Xiaosong Huang, Kewu Wang, Yi Zong
Diabetes, Metabolic Syndrome and Obesity.2023; Volume 16: 779. CrossRef - Managing metabolic diseases: The roles and therapeutic prospects of herb-derived polysaccharides
Xinmei Xu, Lijie Wang, Kun Zhang, Yi Zhang, Gang Fan
Biomedicine & Pharmacotherapy.2023; 161: 114538. CrossRef - Probio-X Relieves Symptoms of Hyperlipidemia by Regulating Patients’ Gut Microbiome, Blood Lipid Metabolism, and Lifestyle Habits
Huan Wang, Cuicui Ma, Yan Li, Lei Zhang, lima A, Chengcong Yang, Feiyan Zhao, Haifeng Han, Dongyang Shang, Fan Yang, Yuying Zhang, Heping Zhang, Zhihong Sun, Ruifang Guo, Yuan Pin Hung
Microbiology Spectrum.2023;[Epub] CrossRef - Sea cucumber sulfated polysaccharides and Lactobacillus gasseri synergistically ameliorate the overweight induced by altered gut microbiota in mice
Zhengqi Liu, Chunqing Ai, Xinping Lin, Xiaoming Guo, Shuang Song, Beiwei Zhu
Food & Function.2023; 14(9): 4106. CrossRef - Supplementation of Lactobacillus plantarum ATCC14917 mitigates non-alcoholic fatty liver disease in high-fat-diet-fed rats
Xingjian Wen, Hejing Liu, Xiaoling Luo, Li Lui, Jiuyu Fan, Yajing Xing, Jia Wang, Xingfang Qiao, Na Li, Guixue Wang
Frontiers in Microbiology.2023;[Epub] CrossRef - Potential herb–drug interactions between anti-COVID-19 drugs and traditional Chinese medicine
Ling Ye, Shicheng Fan, Pengfei Zhao, Chenghua Wu, Menghua Liu, Shuang Hu, Peng Wang, Hongyu Wang, Huichang Bi
Acta Pharmaceutica Sinica B.2023; 13(9): 3598. CrossRef - α‐Lactalbumin Peptide Asp‐Gln‐Trp Ameliorates Hepatic Steatosis and Oxidative Stress in Free Fatty Acids‐Treated HepG2 Cells and High‐Fat Diet‐Induced NAFLD Mice by Activating the PPARα Pathway
Haoran Chen, Yanfeng Ma, Xiaofen Qi, Jianjun Tian, Ying Ma, Tianjiao Niu
Molecular Nutrition & Food Research.2023;[Epub] CrossRef - Lonicerae flos polysaccharides improve nonalcoholic fatty liver disease by activating the adenosine 5′‐monophosphate‐activated protein kinase pathway and reshaping gut microbiota
Chao Han, Zongshuo Li, Ruiying Liu, Zihan Zhao, Yu Wang, Xuli Zuo, Yushi Zhang, Zeyu Geng, Houyu Huang, Xiuzhen Pan, Weidong Li
Journal of the Science of Food and Agriculture.2023; 103(15): 7721. CrossRef - Polysaccharides: The Potential Prebiotics for Metabolic Associated Fatty Liver Disease (MAFLD)
Qin Guo, Yun Li, Xin Dai, Bangmao Wang, Jie Zhang, Hailong Cao
Nutrients.2023; 15(17): 3722. CrossRef - Prebiotics and Probiotics: Therapeutic Tools for Nonalcoholic Fatty Liver Disease
Alejandra Mijangos-Trejo, Natalia Nuño-Lambarri, Varenka Barbero-Becerra, Misael Uribe-Esquivel, Paulina Vidal-Cevallos, Norberto Chávez-Tapia
International Journal of Molecular Sciences.2023; 24(19): 14918. CrossRef - Repair Effect and Mechanism of Electrospinning Nanocomposite Material with Gelatin-Bletilla Striata Gum/Salvia Miltiorrhiza on Orthopedic Refractory Wounds
Geliang Hu, Ming Deng, Yonggang Ma, Jianghua Ming
Journal of Biomedical Nanotechnology.2023; 19(10): 1783. CrossRef -
Characterisation and skin protection activities of polysaccharides from
Schnabelia terniflora
Ying Zhao, Yixian Liu, Huoxiang Zhou, Wei Guo, Weidong Wang, Huiping Chen
Natural Product Research.2023; : 1. CrossRef - Non-alcoholic fatty liver disease risk prediction model and health management strategies for older Chinese adults: a cross-sectional study
Hong Pan, Baocheng Liu, Xin Luo, Xinxin Shen, Jijia Sun, An Zhang
Lipids in Health and Disease.2023;[Epub] CrossRef - Modulatory effects of polysaccharides from plants, marine algae and edible mushrooms on gut microbiota and related health benefits: A review
Henan Zhang, Fuchun Jiang, Jinsong Zhang, Wenhan Wang, Lin Li, Jingkun Yan
International Journal of Biological Macromolecules.2022; 204: 169. CrossRef - Hepatoprotective mechanism of Silybum marianum on nonalcoholic fatty liver disease based on network pharmacology and experimental verification
Guoyan Jiang, Chunhong Sun, Xiaodong Wang, Jie Mei, Chen Li, Honghong Zhan, Yixuan Liao, Yongjun Zhu, Jingxin Mao
Bioengineered.2022; 13(3): 5216. CrossRef - Gut Microbiome in Non-Alcoholic Fatty Liver Disease: From Mechanisms to Therapeutic Role
Haripriya Gupta, Byeong-Hyun Min, Raja Ganesan, Yoseph Asmelash Gebru, Satya Priya Sharma, Eunju Park, Sung-Min Won, Jin-Ju Jeong, Su-Been Lee, Min-Gi Cha, Goo-Hyun Kwon, Min-Kyo Jeong, Ji-Ye Hyun, Jung-A. Eom, Hee-Jin Park, Sang-Jun Yoon, Mi-Ran Choi, Do
Biomedicines.2022; 10(3): 550. CrossRef - Efficacy and Safety of Probiotics Combined With Traditional Chinese Medicine for Ulcerative Colitis: A Systematic Review and Meta-Analysis
Yu Hu, Zhen Ye, Yingqi She, Linzhen Li, Mingquan Wu, Kaihua Qin, Yuzheng Li, Haiqing He, Zhipeng Hu, Maoyi Yang, Fating Lu, Qiaobo Ye
Frontiers in Pharmacology.2022;[Epub] CrossRef - Hepatoprotection of Probiotics Against Non-Alcoholic Fatty Liver Disease in vivo: A Systematic Review
Faezah Sabirin, Siong Meng Lim, Chin Fen Neoh, Kalavathy Ramasamy
Frontiers in Nutrition.2022;[Epub] CrossRef - Salvia miltiorrhiza Bge. (Danshen) in the Treating Non-alcoholic Fatty Liver Disease Based on the Regulator of Metabolic Targets
Jie Liu, Yun Shi, Daiyin Peng, Lei Wang, Nianjun Yu, Guokai Wang, Weidong Chen
Frontiers in Cardiovascular Medicine.2022;[Epub] CrossRef - The Regulatory Roles of Polysaccharides and Ferroptosis-Related Phytochemicals in Liver Diseases
Yijing Ren, Siyue Li, Zixuan Song, Qiuping Luo, Yingying Zhang, Hao Wang
Nutrients.2022; 14(11): 2303. CrossRef - Astaxanthin Alleviates Nonalcoholic Fatty Liver Disease by Regulating the Intestinal Flora and Targeting the AMPK/Nrf2 Signal Axis
Yuhang Li, Juxiong Liu, Bojian Ye, Yueyao Cui, Ruiqi Geng, Shu Liu, Yufei Zhang, Wenjin Guo, Shoupeng Fu
Journal of Agricultural and Food Chemistry.2022; 70(34): 10620. CrossRef - Application of metabolomics in the diagnosis of non-alcoholic fatty liver disease and the treatment of traditional Chinese medicine
Mingmei Shao, Yifei Lu, Hongjiao Xiang, Junmin Wang, Guang Ji, Tao Wu
Frontiers in Pharmacology.2022;[Epub] CrossRef - The chemistry and efficacy benefits of polysaccharides from Atractylodes macrocephala Koidz
Congying Liu, Shengguang Wang, Zedong Xiang, Tong Xu, Mengyuan He, Qing Xue, Huaying Song, Peng Gao, Zhufeng Cong
Frontiers in Pharmacology.2022;[Epub] CrossRef - An Ethnopharmaceutical Study on the Hypolipidemic Formulae in Taiwan Issued by Traditional Chinese Medicine Pharmacies
Min-Han Chi, Jung Chao, Chien-Yu Ko, Shyh-Shyun Huang
Frontiers in Pharmacology.2022;[Epub] CrossRef - Three water soluble polysaccharides with anti-inflammatory activities from Selaginella uncinata (Desv.) Spring
Haochen Hui, Meng Gao, Xuerong Zhao, Lianhong Yin, Lina Xu, Lili Li, Jinyong Peng
International Journal of Biological Macromolecules.2022; 222: 1983. CrossRef - Research Progress on the Therapeutic Effect of Polysaccharides on Non-Alcoholic Fatty Liver Disease through the Regulation of the Gut–Liver Axis
Xiang Chen, Menghan Liu, Jun Tang, Ning Wang, Yibin Feng, Haotian Ma
International Journal of Molecular Sciences.2022; 23(19): 11710. CrossRef - Modulation of gut microbiota by glycyrrhizic acid may contribute to its anti-NAFLD effect in rats fed a high-fat diet
Sai Wang, Xin-Yu Li, Hong-Fang Ji, Liang Shen
Life Sciences.2022; 310: 121110. CrossRef - Structural Alteration of Gut Microbiota During the Amelioration of Chronic Psychological Stress-Aggravated Diabetes-Associated Cognitive Decline by a Traditional Chinese Herbal Formula, ZiBu PiYin Recipe
Wen Zhou, Libin Zhan, Huiying Xu, Lijing Zhang
Journal of Alzheimer's Disease.2022; 90(4): 1465. CrossRef - Bacillus amyloliquefaciens SC06 in the diet improves egg quality of hens by altering intestinal microbiota and the effect is diminished by antimicrobial peptide
Shujie Xu, Fei Wang, Peng Zou, Xiang Li, Qian Jin, Qi Wang, Baikui Wang, Yuanhao Zhou, Li Tang, Dongyou Yu, Weifen Li
Frontiers in Nutrition.2022;[Epub] CrossRef - Effect of stigma maydis polysaccharide on the gut microbiota and transcriptome of VPA induced autism model rats
Xiaolei Yang, Jiyuan Li, Yang Zhou, Ning Zhang, Jicheng Liu
Frontiers in Microbiology.2022;[Epub] CrossRef -
Salvia miltiorrhiza extract may exert an anti-obesity effect in rats with high-fat diet-induced obesity by modulating gut microbiome and lipid metabolism
Zi-Li Ai, Xian Zhang, Wei Ge, You-Bao Zhong, Hai-Yan Wang, Zheng-Yun Zuo, Duan-Yong Liu
World Journal of Gastroenterology.2022; 28(43): 6131. CrossRef - Dysregulated hepatic lipid metabolism and gut microbiota associated with early-stage NAFLD in ASPP2-deficiency mice
Fang Xie, Hang-fei Xu, Jing Zhang, Xiao-ni Liu, Bu-xin Kou, Meng-yin Cai, Jing Wu, Jin-ling Dong, Qing-hua Meng, Yi Wang, Dexi Chen, Yang Zhang
Frontiers in Immunology.2022;[Epub] CrossRef - Structural elucidation and anti-nonalcoholic fatty liver disease activity of Polygonatum cyrtonema Hua polysaccharide
Wei Liu, Taili Shao, Lei Tian, Zhengrui Ren, Lan Gao, Zhiyan Tang, Zheng Fang, Pingchuan Yuan, Chunyan Liu, Jikun Li, Guodong Wang, Jun Han
Food & Function.2022; 13(24): 12883. CrossRef - The protective effects of sulforaphane on high-fat diet-induced metabolic associated fatty liver disease in mice via mediating the FXR/LXRα pathway
Shaotong Ma, Xinyi Pang, Shuhua Tian, Jing Sun, Qiaobin Hu, Xiangfei Li, Yingjian Lu
Food & Function.2022; 13(24): 12966. CrossRef - Suppression of obesity and inflammation by polysaccharide from sporoderm-broken spore of Ganoderma lucidum via gut microbiota regulation
Tingting Sang, Chengjie Guo, Dandan Guo, Jianjun Wu, Yujie Wang, Ying Wang, Jiajun Chen, Chaojie Chen, Kaikai Wu, Kun Na, Kang Li, Liu Fang, Cuiling Guo, Xingya Wang
Carbohydrate Polymers.2021; 256: 117594. CrossRef - Annual review of LSD1/KDM1A inhibitors in 2020
Dong-Jun Fu, Jun Li, Bin Yu
European Journal of Medicinal Chemistry.2021; 214: 113254. CrossRef - Diet-Regulating Microbiota and Host Immune System in Liver Disease
Jung A Eom, Goo Hyun Kwon, Na Yeon Kim, Eun Ju Park, Sung Min Won, Jin Ju Jeong, Ganesan Raja, Haripriya Gupta, Yoseph Asmelash Gebru, Satyapriya Sharma, Ye Rin Choi, Hyeong Seop Kim, Sang Jun Yoon, Ji Ye Hyun, Min Kyo Jeong, Hee Jin Park, Byeong Hyun Min
International Journal of Molecular Sciences.2021; 22(12): 6326. CrossRef - Lactobacillus johnsonii BS15 combined with abdominal massage on intestinal permeability in rats with nonalcoholic fatty liver and cell biofilm repair
Wei Zhang, Huanan Li, Na Zhao, Xiongfei Luo, Siwen Liu, an Bao, Yingying Chen, Haiteng Wang, Junshi Wang, Jingui Wang
Bioengineered.2021; 12(1): 6354. CrossRef - Effects and Mechanisms of Probiotics, Prebiotics, Synbiotics, and Postbiotics on Metabolic Diseases Targeting Gut Microbiota: A Narrative Review
Hang-Yu Li, Dan-Dan Zhou, Ren-You Gan, Si-Yu Huang, Cai-Ning Zhao, Ao Shang, Xiao-Yu Xu, Hua-Bin Li
Nutrients.2021; 13(9): 3211. CrossRef - Correlation Analysis of Huayu Tongmai Decoction Intervention and Prognosis Indexes of Patients with Carotid Atherosclerosis
Guangqing Cheng, Xiaoni Yan, Fengmeng Wang, Chao Chen, Muhammad Wasim Khan
Evidence-Based Complementary and Alternative Medicine.2021; 2021: 1. CrossRef - Targeting the gut microbiota by Asian and Western dietary constituents: a new avenue for diabetes
Abdul Rahman Conteh, Ruixue Huang
Toxicology Research.2020; 9(4): 569. CrossRef - MiR-455 targeting SOCS3 improve liver lipid disorders in diabetic mice
Shu Fang, Jie Feng, Hongbin Zhang, Ping Li, Yudan Zhang, Yanmei Zeng, Yingying Cai, Xiaochun Lin, Yaoming Xue, Meiping Guan
Adipocyte.2020; 9(1): 179. CrossRef - Chinese Medicinal Herbs Targeting the Gut–Liver Axis and Adipose Tissue–Liver Axis for Non-Alcoholic Fatty Liver Disease Treatments: The Ancient Wisdom and Modern Science
Shuwei Zhang, Yui-Tung Wong, Ka-Yu Tang, Hiu-Yee Kwan, Tao Su
Frontiers in Endocrinology.2020;[Epub] CrossRef - Probiyotiklerin Kolon Mikrobiyotasına Etkileri: Güncel Çalışmalar
Çağlar GÖKIRMAKLI, Zeynep SEYDİM
Journal of Biotechnology and Strategic Health Research.2020; 4(3): 212. CrossRef
 PubReader
PubReader Cite
Cite