- Current
- Browse
- Collections
-
For contributors
- For Authors
- Instructions to authors
- Article processing charge
- e-submission
- For Reviewers
- Instructions for reviewers
- How to become a reviewer
- Best reviewers
- For Readers
- Readership
- Subscription
- Permission guidelines
- About
- Editorial policy
Articles
- Page Path
- HOME > Diabetes Metab J > Volume 35(5); 2011 > Article
-
Original ArticleDietary Oleate Has Beneficial Effects on Every Step of Non-Alcoholic Fatty Liver Disease Progression in a Methionine- and Choline-Deficient Diet-Fed Animal Model
- Ji Young Lee1, Jae Hoon Moon2, Jong Suk Park2, Byung-Wan Lee2, Eun Seok Kang2, Chul Woo Ahn2, Hyun Chul Lee2, Bong Soo Cha1,2
-
Diabetes & Metabolism Journal 2011;35(5):489-496.
DOI: https://doi.org/10.4093/dmj.2011.35.5.489
Published online: October 31, 2011
1Brain Korea 21 Project for Medical Science, Yonsei University College of Medicine, Seoul, Korea.
2Department of Internal Medicine, Yonsei University College of Medicine, Seoul, Korea.
- Corresponding author: Bong Soo Cha. Department of Internal Medicine, Yonsei University College of Medicine, 250 Seongsanno, Seodaemun-gu, Seoul 120-752, Korea. bscha@yuhs.ac
- *Ji Young Lee and Jae Hoon Moon contributed equally to this work as first authors.
• Received: February 11, 2011 • Accepted: April 11, 2011
Copyright © 2011 Korean Diabetes Association
This is an Open Access article distributed under the terms of the Creative Commons Attribution Non-Commercial License (http://creativecommons.org/licenses/by-nc/3.0/) which permits unrestricted non-commercial use, distribution, and reproduction in any medium, provided the original work is properly cited.


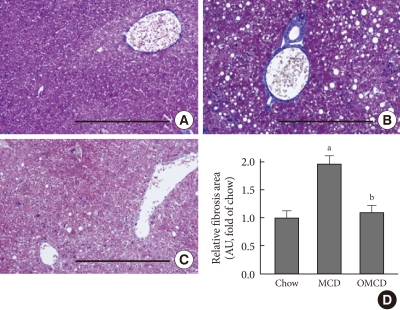

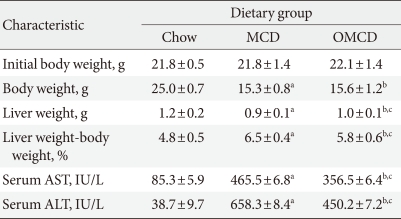
Figure & Data
References
Citations
Citations to this article as recorded by 
- Bidirectional association between NAFLD and gallstone disease: a systematic review and meta-analysis of observational studies
Shengying Gu, Shanshan Hu, Shuowen Wang, Chendong Qi, Chenyang Shi, Guorong Fan
Expert Review of Gastroenterology & Hepatology.2023; 17(3): 283. CrossRef - The Effect of Bioactive Aliment Compounds and Micronutrients on Non-Alcoholic Fatty Liver Disease
Camelia Munteanu, Betty Schwartz
Antioxidants.2023; 12(4): 903. CrossRef - Single‐cell transcriptomics stratifies organoid models of metabolic dysfunction‐associated steatotic liver disease
Anja Hess, Stefan D Gentile, Amel Ben Saad, Raza‐Ur Rahman, Tim Habboub, Daniel S Pratt, Alan C Mullen
The EMBO Journal.2023;[Epub] CrossRef - Histopathological Examination of the Effects of Tocilizumab and Dexamethasone on the Liver in Rats of Oleic Acid induced Acute Lung Injury
Funda TERZİ, Hüseyin Serkan EROL
Balıkesır Health Sciences Journal.2022;[Epub] CrossRef - Identifying Lipid Metabolites Influenced by Oleic Acid Administration Using High-Performance Liquid Chromatography–Mass Spectrometry-Based Lipidomics
Chao Xu, Dan Song, Askild L. Holck, Youyou Zhou, Rong Liu
ACS Omega.2020; 5(20): 11314. CrossRef - Causative and Sanative dynamicity of ChREBP in Hepato-Metabolic disorders
P. Vineeth Daniel, Prosenjit Mondal
European Journal of Cell Biology.2020; 99(8): 151128. CrossRef - PPARδ attenuates hepatic steatosis through autophagy-mediated fatty acid oxidation
Lei Tong, Long Wang, Shuangshuang Yao, Lina Jin, Jian Yang, Yifei Zhang, Guang Ning, Zhiguo Zhang
Cell Death & Disease.2019;[Epub] CrossRef - Butyrate Protects Mice Against Methionine–Choline-Deficient Diet-Induced Non-alcoholic Steatohepatitis by Improving Gut Barrier Function, Attenuating Inflammation and Reducing Endotoxin Levels
Jianzhong Ye, Longxian Lv, Wenrui Wu, Yating Li, Ding Shi, Daiqiong Fang, Feifei Guo, Huiyong Jiang, Ren Yan, Wanchun Ye, Lanjuan Li
Frontiers in Microbiology.2018;[Epub] CrossRef - Olive oil combined with Lycium barbarum polysaccharides attenuates liver apoptosis and inflammation induced by carbon tetrachloride in rats
Yun-Yun Chiang, Jane C.-J. Chao
Journal of Functional Foods.2018; 48: 329. CrossRef - Dietary oleic acid regulates hepatic lipogenesis through a liver X receptor-dependent signaling
Simon Ducheix, Alexandra Montagner, Arnaud Polizzi, Frédéric Lasserre, Marion Régnier, Alice Marmugi, Fadila Benhamed, Justine Bertrand-Michel, Laila Mselli-Lakhal, Nicolas Loiseau, Pascal G. Martin, Jean-Marc Lobaccaro, Laurent Ferrier, Catherine Postic,
PLOS ONE.2017; 12(7): e0181393. CrossRef - Is hepatic lipogenesis fundamental for NAFLD/NASH? A focus on the nuclear receptor coactivator PGC-1β
Simon Ducheix, Maria Carmela Vegliante, Gaetano Villani, Nicola Napoli, Carlo Sabbà, Antonio Moschetta
Cellular and Molecular Life Sciences.2016; 73(20): 3809. CrossRef - Metformin alleviates hepatosteatosis by restoring SIRT1-mediated autophagy induction via an AMP-activated protein kinase-independent pathway
Young Mi Song, Yong-ho Lee, Ji-Won Kim, Dong-Sik Ham, Eun-Seok Kang, Bong Soo Cha, Hyun Chul Lee, Byung-Wan Lee
Autophagy.2015; 11(1): 46. CrossRef - Adipokines and proinflammatory cytokines, the key mediators in the pathogenesis of nonalcoholic fatty liver disease
Sanja Stojsavljević
World Journal of Gastroenterology.2014; 20(48): 18070. CrossRef - Microglial Cell Activation Increases Saturated and Decreases Monounsaturated Fatty Acid Content, but Both Lipid Species are Proinflammatory
Emily B. Button, Andrew S. Mitchell, Marcia M. Domingos, Jessica H.‐J. Chung, Ryan M. Bradley, Ashkan Hashemi, Phillip M. Marvyn, Ashley C. Patterson, Ken D. Stark, Joe Quadrilatero, Robin E. Duncan
Lipids.2014; 49(4): 305. CrossRef - Modeling progressive non-alcoholic fatty liver disease in the laboratory mouse
Jesse D. Riordan, Joseph H. Nadeau
Mammalian Genome.2014; 25(9-10): 473. CrossRef - Rapid chromatographic method to decipher distinct alterations in lipid classes in NAFLD/NASH
Stephan Laggai, Yvette Simon, Theo Ranssweiler, Alexandra K Kiemer, Sonja M Kessler
World Journal of Hepatology.2013; 5(10): 558. CrossRef - Dimethyl sulfoxide reduces hepatocellular lipid accumulation through autophagy induction
Young Mi Song, Sun-Ok Song, Yong-Keun Jung, Eun-Seok Kang, Bong Soo Cha, Hyun Chul Lee, Byung-Wan Lee
Autophagy.2012; 8(7): 1085. CrossRef
 PubReader
PubReader Cite
Cite