- Current
- Browse
- Collections
-
For contributors
- For Authors
- Instructions to authors
- Article processing charge
- e-submission
- For Reviewers
- Instructions for reviewers
- How to become a reviewer
- Best reviewers
- For Readers
- Readership
- Subscription
- Permission guidelines
- About
- Editorial policy
Articles
- Page Path
- HOME > Diabetes Metab J > Volume 42(6); 2018 > Article
-
ReviewOthers A Journey to Understand Glucose Homeostasis: Starting from Rat Glucose Transporter Type 2 Promoter Cloning to Hyperglycemia
-
Yong Ho Ahn

-
Diabetes & Metabolism Journal 2018;42(6):465-471.
DOI: https://doi.org/10.4093/dmj.2018.0116
Published online: November 2, 2018
Department of Biochemistry and Molecular Biology, Integrated Genomic Research Center for Metabolic Regulation, Yonsei University College of Medicine, Seoul, Korea.
- Corresponding author: Ahn Yong Ho. Department of Biochemistry and Molecular Biology, Integrated Genomic Research Center for Metabolic Regulation, Yonsei University College of Medicine, 50 Yonsei-ro, Seodaemun-gu, Seoul 03722, Korea. yha111@yuhs.ac
• Received: July 30, 2018 • Accepted: August 8, 2018
Copyright © 2018 Korean Diabetes Association
This is an Open Access article distributed under the terms of the Creative Commons Attribution Non-Commercial License (http://creativecommons.org/licenses/by-nc/4.0/) which permits unrestricted non-commercial use, distribution, and reproduction in any medium, provided the original work is properly cited.
- ABSTRACT
- INTRODUCTION
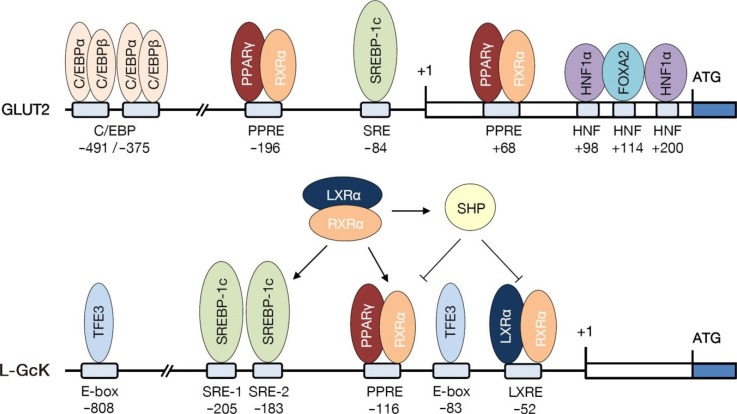
- STARTING FROM RAT GLUT2 PROMOTER
- EXPANSION TO HUMAN GLUT2 PROMOTER
- NUCLEAR RECEPTOR AND GLUCOSE SENSOR REGULATION IN LIVER AND β-CELLS
- SREBP-1c AND GLUCOSE METABOLISM
- ROLE OF PPARα AND PROLACTIN REGULATORY ELEMENT-BINDING ON THE GLUCONEOGENESIS
- ROLE OF PROTEIN INTERACTIONS AND GLUCOSE HOMEOSTASIS
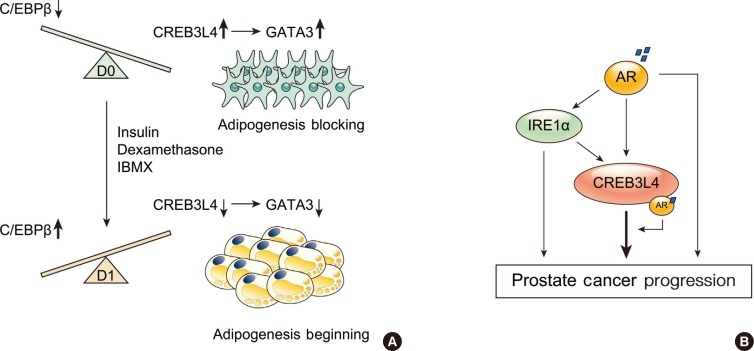
- ROLE OF CREB3L4 ON THE ADIPOGENESIS
- SUMMARY
- ACKNOWLEDGMENTS
- NOTES
- REFERENCES

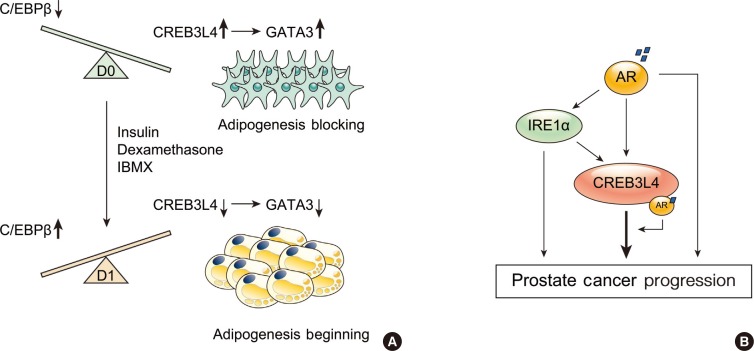
Figure & Data
References
Citations
Citations to this article as recorded by 
- Physiological functions of glucose transporter-2: From cell physiology to links with diabetes mellitus
Zhean Shen, Yingze Hou, Guo Zhao, Libi Tan, Jili Chen, Ziqi Dong, Chunxiao Ni, Longying Pei
Heliyon.2024; 10(3): e25459. CrossRef - Estimation and implications of the genetic architecture of fasting and non-fasting blood glucose
Zhen Qiao, Julia Sidorenko, Joana A. Revez, Angli Xue, Xueling Lu, Katri Pärna, Harold Snieder, Peter M. Visscher, Naomi R. Wray, Loic Yengo
Nature Communications.2023;[Epub] CrossRef - Umbilical Cord-Mesenchymal Stem Cell-Conditioned Medium Improves Insulin Resistance in C2C12 Cell
Kyung-Soo Kim, Yeon Kyung Choi, Mi Jin Kim, Jung Wook Hwang, Kyunghoon Min, Sang Youn Jung, Soo-Kyung Kim, Yong-Soo Choi, Yong-Wook Cho
Diabetes & Metabolism Journal.2021; 45(2): 260. CrossRef - Aging-related modifications to G protein-coupled receptor signaling diversity
Jaana van Gastel, Hanne Leysen, Jan Boddaert, Laura vangenechten, Louis M. Luttrell, Bronwen Martin, Stuart Maudsley
Pharmacology & Therapeutics.2021; 223: 107793. CrossRef - Glucose transporters in the small intestine in health and disease
Hermann Koepsell
Pflügers Archiv - European Journal of Physiology.2020; 472(9): 1207. CrossRef
 PubReader
PubReader Cite
Cite