Abnormal Responses in Cognitive Impulsivity Circuits Are Associated with Glycosylated Hemoglobin Trajectories in Type 1 Diabetes Mellitus and Impaired Metabolic Control
Article information

Abstract
Background
Risky health decisions and impulse control profiles may impact on metabolic control in type 1 diabetes mellitus (T1DM). We hypothesize that the neural correlates of cognitive impulsivity and decision-making in T1DM relate to metabolic control trajectories.
Methods
We combined functional magnetic resonance imaging (fMRI), measures of metabolic trajectories (glycosylated hemoglobin [HbA1c] over multiple time points) and behavioral assessment using a cognitive impulsivity paradigm, the Balloon Analogue Risk Task (BART), in 50 participants (25 T1DM and 25 controls).
Results
Behavioral results showed that T1DM participants followed a rigid conservative risk strategy along the iterative game. Imaging group comparisons showed that patients showed larger activation of reward related, limbic regions (nucleus accumbens, amygdala) and insula (interoceptive saliency network) in initial game stages. Upon game completion differences emerged in relation to error monitoring (anterior cingulate cortex [ACC]) and inhibitory control (inferior frontal gyrus). Importantly, activity in the saliency network (ACC and insula), which monitors interoceptive states, was related with metabolic trajectories, which was also found for limbic/reward networks. Parietal and posterior cingulate regions activated both in controls and patients with adaptive decision-making, and positively associated with metabolic trajectories.
Conclusion
We found triple converging evidence when comparing metabolic trajectories, patients versus controls or risk averse (non-learners) versus patients who learned by trial and error. Dopaminergic reward and saliency (interoceptive and error monitoring) circuits show a tight link with impaired metabolic trajectories and cognitive impulsivity in T1DM. Activity in parietal and posterior cingulate are associated with adaptive trajectories. This link between reward-saliency-inhibition circuits suggests novel strategies for patient management.
INTRODUCTION
Type 1 diabetes mellitus (T1DM) patients are faced with systematic habit-related daily decision-making to achieve metabolic control and to avoid future uncertain health complications. It is expected that they monitor blood glucose, follow a diet, calculate carbohydrates for each mealtime while being vigilant to body signals of glycemic status, which they learn by routine. These disease-related tasks are relevant to achieve efficient self-management. Effective decision-making in daily life routines implies control of impulsivity towards unhealthy “rewards.” From a psychobiological perspective, impulsivity is characterized by failure in inhibiting a potentially risky impulse. From a cognitive viewpoint, impulsivity represents the inability to inhibit behavioral impulses with impact on social and personal functioning.
Adaptive decision-making is related to such inhibitory self-control. Studies with T1DM people reported that engaging in impulsive behaviors without forethought and prematurely responding to stimuli. Moreover it is important to study the ability of patients to anticipate long term health “rewards.” This ability to inhibit non-adaptive responses taking into account long term benefits is called delay discounting, and justifies the need to study cognitive impulsivity using appropriate tasks. Impulsive patterns of behavior and related personality traits are mediators between self-management and glycemic control [1,2]. We have previously shown that in T1DM behavioral risk-taking phenotypes can be predicted by the dynamics of glycosylated hemoglobin (HbA1c) [1]. We adopted a strategy of using a battery of self-reported and real-world risk behaviors (concerning multiple domains of risk-taking behavior) and an experimental cognitive impulsivity task the Balloon Analogue Risk Task (BART), which is well suited to study cognitive impulsivity [3]. Such tasks involve trading off positive and negative rewards and are therefore also known as risk-taking tasks. Using this behavioral approach we found that behavioral risk-taking endophenotypes relate to the division between improving metabolic control versus deteriorating states [1], providing the motivation of the current study.
This previous study [1] did not take just a single measure of HbA1c, but multiple measures along time. The fact that we could observe dynamics and not a single snapshot is very important and allowed to, using K-means and two-step cluster analysis, to identify a two-cluster solution providing information of distinct decision profiles (concerning multiple domains of impulsive risk-taking behavior) which match the biological partition, based on the division between stable or improving metabolic control [1]. This sets the ground for the current study which aims to identify the neural correlates of impulsive behavior in T1DM and this biological partition.
It is important to note that the study of cognitive impulsivity is directly related to adaptive decision-making and reward evaluation. People with T1DM must evaluate cost/benefits, to anticipate risk/reward outcomes and to assess delay in time for long term healthy rewards (delay discounting). In general, this requires self-regulation skills which help to learn and adaptively respond to dynamic contexts. For example, maladaptive risk-taking is related to impulsive patterns of behavior and is relevant in chronic disorders [4,5]. It has been suggested that motivated goal-directed behavior with self-relevant consequences is related to brain systems related to impulsivity and inhibitory control in prefrontal cortex, and reward/limbic regions such as the nucleus accumbens (NAcc) and amygdala [6-8].
Neuroimaging studies using the BART, the task we chose to study cognitive impulsivity in T1DM based on previous findings [1], reported cortical and subcortical regions associated with risky decisions and uncertainty [9-11], suggesting that this task is appropriate to study cognitive impulsivity in clinical populations [3]. These studies suggest an important role for the anterior cingulate cortex (ACC) and insula. Both of these regions belong to the saliency network, which is recruited in response to environments crowded with uncertainty [12]. “Cash out” events (reflecting the decision to keep the present accumulated reward and leave safely the game), also interpreted in previous BART studies as related to “win” [13] and “relief ” [10], recruited dopaminergic reward related regions, while “explosions” (reflecting sudden loss of reward, because of keeping the impulse to stay in the game) recruited regions involved in saliency and interoceptive processing [14,15]. Decision-making requiring impulse control is linked to several interrelated networks relevant to disease state monitoring. First, the salience network (insula and dorsal ACC), is relevant to the integration of emotional, cognitive, and behavioral self-regulation [16]. Second, the limbic system which is pivotal in emotional regulation. The limbic and reward/motivational systems are strongly connected [17]. Thus, the reward/motivational system is associated with pathways that share limbic structures [18,19]. Third, cognitive control of impulsivity is also related the inferior frontal gyrus (IFG) which is pivotal in response inhibition [20]. When studying decision-making it is important to take into account subjective value, of rewards/outcomes and neural processes that weight risk and regulate ultimate decision [21].
The present study combines functional magnetic resonance imaging with temporal measures of HbA1c and behavioral assessment using the BART, an experimental task to test impulse control under ambiguity which can predict dynamics of metabolic control levels in T1DM [1]. We therefore used BART to examine the neural correlates of impulsivity, which in the case of diabetes comprises the impulsivity features that patients need to regulate for successful metabolic control. Using this task during functional magnetic resonance imaging (fMRI), we aimed to understand the neural mechanisms of risky decision-making and impulse control since the BART task allows (1) to simulate a tension between reward seeking and loss aversion, involving cognitive control of impulse and motivational processes; (2) to investigate naturalistic risk-taking behaviors requiring to weight short and long term rewards. This sequential decision-making task is defined by an escalating risk with an increased reward on a trial-by-trial basis. It allows for error monitoring through successive “balloon” inflations (accumulating reward) and unexpected explosions (losing all), leading to potential shifts between averse and seeking choices, helping to define a dynamic impulse control behavioral profile after iterative decisions.
The task involved four periods. (1) Before iterative decision-making: This is named as the initial period (first balloon series out of a total of 30 balloon series) or unlearned task performance occurring with the first task administration, called guessing. (2) End of game, after iterative decision-making: This period is also described as the final period (last balloon series, the 30th) or learned adaptive impulse control after iterations. (3) Appetitive outcome (cash out decisions related to taking immediate rewards, while inhibiting the impulse to continue). (4) Aversive outcome (the explosion outcome related to putative risk-taking choices that led to negative outcomes).
Along this line, this study aims to identify whether impulsivity phenotypes related to risk averse versus risk seeking behavior, whose profiles come from task performance, present distinct neural phenotypes. Risk aversiveness is defined here as preference for choosing cash out decisions than to risk the balloon explosion. It means a preference for lower and immediate rewards than large and less probable amounts of money.
Importantly, this study set out to find out the correlation between dynamic biological worsening over time and risk-taking behavior in the four periods previously described by relating brain responses with the progression of individual values of HbA1c over time across multiple time points: (1) before iterative decision-making (prior to trial and error learning); (2) after iterative decision-making (after trial and error learning); (3) appetitive outcomes (cashing); and (4) aversive outcomes (balloon explosions and full loss).
We designed our BART-fMRI studies to test the hypothesis that T1DM will present an overactivation of brain regions related to impulse control, reward and saliency processing (which directly relates to interoception).
METHODS
Participants
Written consent was obtained from all participants and the study was approved by the Ethics Committee (CE-002/2014) of the Faculty of Medicine of University of Coimbra, in accordance with the Declaration of Helsinki. All participants had normal or corrected-to-normal vision, were right-handed and had no history of neurological or psychiatric disorders. Each participant met standard safety requirements for entry into the magnetic resonance imaging scanner. They were paid depending on the money they earned during the experimental task to render it ecologically valid.
We recruited 50 adults aged 22 to 55 years (Supplementary Fig. 1 for study workflow). Experiments were conducted between October 2015 and August 2017. Clinical analyses to controls were made at University Hospital of Coimbra (CHUC) to assure that no one had diagnosis of diabetes mellitus, diagnosed according to the current World Health Organization criteria. Twenty-five of them were diagnosed with T1DM (age: mean±standard deviation [SD], 38.72±10.38; age range, 22 to 55; 11 males and 14 females; HbA1c, 7.86±1.29; HbA1c range, 5.9 to 11.6) at time of scanning. The remaining 25 were matched healthy individuals (age, mean±SD, 35.08±8.77; age range, 24 to 55, 10 males and 15 females; HbA1c, 4.98±0.25; HbA1c range, 4.5 to 5.7). Groups were matched according to gender, age, civil state, and household members (Table 1).

Demographic characteristics, cognitive results, and self-reported measures in T1DM and healthy participants (n=50)
Procedures
Self-reported individual risk
Individual self-reported real-world risk profile was accessed by a comprehensive battery of questions made for this purpose covering three levels: (1) context (domain-specific risk-taking [DOSPERT] scale [22], Portuguese translation [23], allowed to achieve individual perception of risk-taking in health and financial contexts); (2) time perspective of risk (general past and present risk-taking questions catch out the influence of time in risk profile perception because participants were asked to compare the same type of risk in different time points [10 years ago and present time]); and finally, (3) capacity to delay reward (temporal discounting—the ability to weight short vs. long term benefits) (Intertemporal choice questions were used to access the preference for delayed over immediate rewards). Here, participants chose between three options: a smaller earlier reward (SS), an intermediate (II) or larger longer reward (LL). They had three thematic decision challenges: financial, health, and specific health context (diabetes). Risk related constructs as impulsivity, personality and eating behavior were evaluated by Behavior Impulsivity Scale-11 (BIS-11; translated by [24]) and validated for the Portuguese population by [25], Eysenck Personality Questionnaire (EPQ, Portuguese version) [26] and Portuguese validation of Dutch Eating Behavior Questionnaire (DEBQ) [27,28]. It evaluates three types of eating styles such as restrained (avoid eating), external (eating motivated by smell or visual attractiveness), and emotional (eating in response to emotions).
Balloon Analogue Risk Task
Participants performed a version of the BART developed based on the original implementation of the BART for fMRI [13]. Before starting the scanning session, the task was explained using a static template accompanied by specific instructions (Supplementary material). The money accumulated throughout the experiment was paid in cash to assure participants’ engagement during the task. BART behavioral measurements were compared to characterize the groups (Fig. 1)
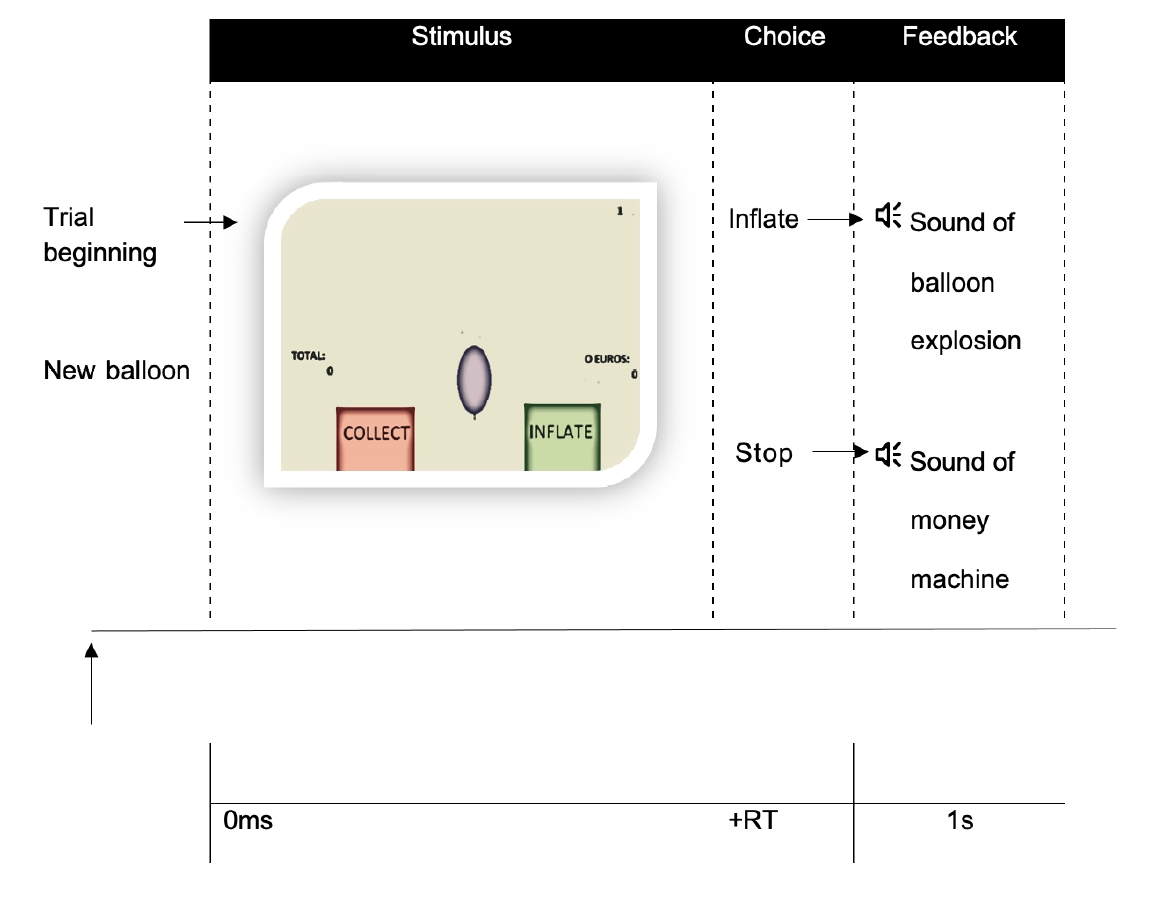
Schematic diagram for a trial sequence in the Balloon Analogue Risk Task (BART) at functional magnetic resonance imaging. For each balloon (stimulus), participants had to decide (choice) inflate the balloon or collet the money earned. The consequence of that choice (feedback) was revealed by a sound of balloon explosion (if the balloon exploded) or a sound of a money machine (if they collected the money). Participants were presented with 30 balloon series, one balloon at a time in the center of the screen. To make decisions, participants used a button-press to click to inflate (right button) or to click to stop inflate and save the money (left button). Participants do not know in advance the exact probability of explosion nor the maximum number of inflations. RT, reaction time.
Behavior and fMRI data analysis
Behavioral data were analysed using IBM SPSS Statistic version 24 (IBM Co., Armonk, NY, USA). Descriptive statistics are reported as mean±SD. Prior to analysis raw data were examined for normality by Shapiro-Wilks goodness-of-fit test. Null-hypothesis statistical tests were evaluated according to an alpha value of P=0.05.
For each subject, functional images were pre-processed using BrainVoyager QX software (https://www.brainvoyager.com) and consisted of slice scan time correction, High temporal filtering and three-dimensional motion correction via realignment. The functional image was co-registered to the anatomical image. Before statistical analysis, images were normalized to the Talairach space and were then spatially smoothed using an 8 mm full-width-half-maximum Gaussian kernel. Because we would like to define predictors that are closely spaced, we applied a deconvolution analysis for each subject to separate the overlapping responses to four conditions or single data points: first balloon in series, last balloon in series, cash out decision and explode outcome. We performed a group analysis and run a deconvolution analysis on task events. The predictors were defined based on the selection of 10 data points (each condition with 10 “sticky” predictors of interest, from D0 up to D9). In Generalized Linear Model (GLM), we specified the overlay contrasts as D0–D3 (minus), D4–D6 (plus), and D7–D9 (minus). In the deconvolution analysis we created 10 shifted predictors (D0–D9 data points, where the repetition time [TR]=2 seconds) for each analyzed condition, ensuring that we cover the whole hemodynamic response of each of the four conditions (at the end, we have 40 predictors).
Thereafter, we balanced and created separate maps for each subject, comparing the two groups (T1DM and healthy participants) for each condition. All contrast maps (beta maps) within a between subject analysis were calculated to identify the neural substrates associated with each condition to both groups. To correct for multiple comparisons a statistical threshold of P<0.05 was fixed and a minimum cluster size threshold was estimated using Monte Carlo simulations (1,000 iterations). The number of contiguous voxels considered as the minimum cluster extension for each map is presented with the statistical maps. We conducted analysis of fMRI data separately for the first balloon (prior to learning) and the last balloon (after learning through iteration). We generated statistical maps contrasting T1DM and healthy groups during first balloon and during last balloon of the experiment (to assess task learning). We repetead the same procedure to generate statistical maps contrastring risk averse and risk seeking subgroups within patients. These subgroups were planned according to a median cut-off of number of inflations (see below). Thereafter, we made the same procedure between group analysis (T1DM vs. healthy participants; and risk averse vs. risk seeking groups) with two different predictors: cash out decision and explode outcome. Finally, a linear function was adjusted to the progression individual values of HbA1c over time. It means, that multiple HbA1c values for each patient were obtained from individual healthcare process since they were diagnosed with T1DM. The regressor calculated to each patient was used to define successful metabolic control (negative slope, i.e., decreasing HbA1c values over the time) and difficult metabolic control (positive slope, i.e., increasing HbA1c values over the time). Patients were assessed over an interval of at least 2 years and up to 8 years (with visits every 6 months), allowing to obtain rich dynamic information. Since we measured multiple time points we were able to determine positive or negative slopes of evolution of metabolic control which were then correlated with conditions of interest (namely neural responses to cash out or explosion events). As described in a previous publication (1), HbA1c trajetories were obtained by retrieving dynamic values over time. A stable HbA1c trend with an increased range of HbA1c may indeed indicate poor metabolic control and this is captured by a slope measure. Frequency of Hypoglycemia was measured and we found an association with metabolic control (χ2 (1)=7.94, P=0.006, d=0.62). The group with less metabolic control (1) was more strongly associated with the presence of hypoglycemia.
RESULTS
Behavioral risk measures between T1DM and Healthy participants
Self-report individual risk
We first report the results concerning, impulsive behavior profiles in general contexts and also in the health context of diabetes. Mann-Whitney tests revealed that T1DM and healthy groups did not differ in self-reported measures of risk-taking, except for delay rewards. Accordingly, healthy subjects tended to opt for delayed over immediate rewards in all contexts, whereas T1DM opted only for delay rewards when the choice situation is related to diabetes—preference for current management (more daily pricks) in order to have less future complications (blindness). This suggests that their decision profiles are in general maladaptive preferring short term earlier rewards except in the health context of diabetes (possibly reflecting health education). Regarding eating behavior, people with T1DM reported lower scores in external and restrained eating behavior as compared to the healthy group (Table 1).
Balloon Analogue Risk Task
Table 2 shows behavioral results acquired during the fMRI experiment. Nonparametric independent sample tests revealed significant differences between groups. Participants in the T1DM group chose more cash out than the control group and the number of inflations for the first balloon is similar to the last balloon (t(24)=–2.64, P=0.794). For healthy subjects the opposite pattern is found: the number of inflations in the last balloon is higher than in the first balloon, converging to a change in risk profile (t(24)=–2.53, P=0.018) (Supplementary Fig. 2).
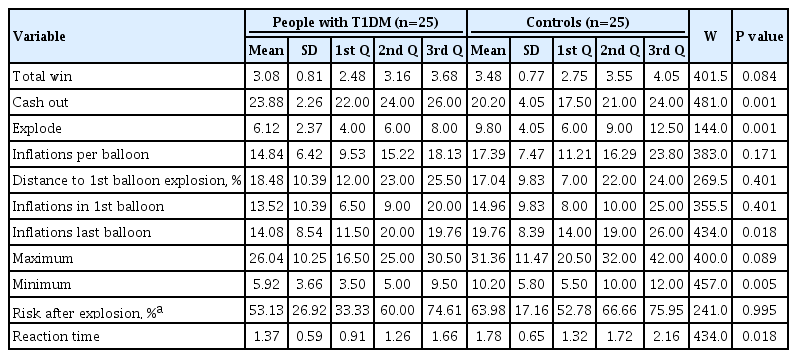
Behavioral results on BART task (n=50), T1DM, and healthy groups
Behavioral risk measures between risk averse and risk seeking groups within T1DM
To address the relevance of risk-taking profiles, we performed a planned subgroup analysis. Considering T1DM patients (n=25), we divided them in two groups according to the performance on the risk task. We used a median cut-off point on 20 pumps for each balloon (average of pumps for the group). For each participant, we recorded the frequency of number of pumps for each balloon considering all participants in all trials (30 balloon series). Participants’ results were ordered and then split according to the median cut-off point (less frequent pumps-risk averse group; high frequency pumps-risk averse group). Twenty pumps refers to the median of pumps for the group (median or Q50). This yielded a group with 11 patients with risk averse profile (mean age, 36.18±10.34; age range, 22 to 47; 6 males and 5 females) and a group with 14 patients with risk-taking profile (mean age, 40.71±10.57; age range, 22 to 55; 5 males and 9 females). Groups were matched according to all sociodemographic variables.
Self-reported risk measure
Between group analysis showed that T1DM with risk averse performance scored higher on lack of planning (U=38.5, P=0.034) and scored lower on health risk perception (U=119.5, P=0.018) than T1DM with a more adaptive risk-taking (searching choice space) performance. Risk averse participants tended to score higher on inhibitory control (U=41.5, P=0.051).
Balloon Analogue Risk Task
As expected, both groups differ in almost all behavioral variables of BART experimental task (Supplementary Table 1).
Neuroimaging results
T1DM vs. healthy groups
1) First balloon series
During the first balloon series (beginning of game), participants were deciding to inflate under complete uncertainty and ambiguity. Differential activations in patients included clusters in emotion and reward related regions such as right amygdala, ventral (accumbens), and dorsal striatum (putamen, caudate). Differential patient activations in saliency network regions involved in decision under ambiguity (insula) or inhibitory control (bilateral IFG) were also found (Fig. 2).
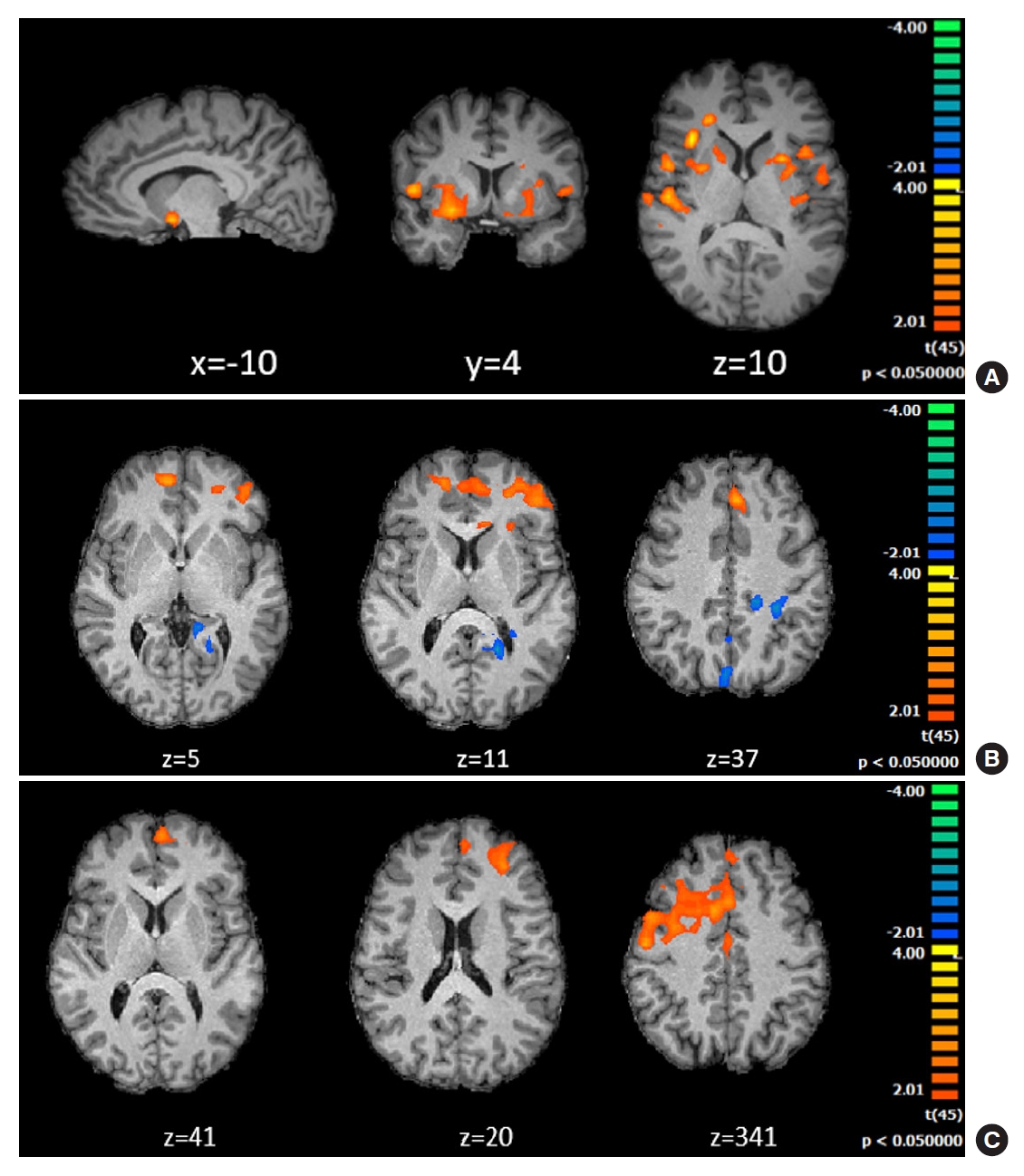
Group differences between the type 1 diabetes mellitus (T1DM) and control group during first balloon (A), last balloon (B, end of game) and “explode” (C) outcome condition of the impulse control Balloon Analogue Risk Task (BART) task. (A) Differential activations in limbic, reward, saliency and inhibitory control regions: right amygdala, ventral, and dorsal striatum (nucleus accumbens, putamen, and caudate), insula (saliency network) and inferior frontal gyrus (minimum cluster size 78 voxels, for details see methods). (B) Last balloon condition (after trial and error learning in iterative decision-making). T1DM showed higher activity in regions related to error monitoring, in anterior saliency network regions such as bilateral anterior cingulate cortex (ACC; BA32, BA24) and frontal regions (9, 10, 8, 45). Patients revealed decreased blood oxygen level dependent (BOLD) activity (blue) in posterior cingulate cortex (BA23, 30, 31) (minimum cluster size 95 voxels). (C) Group differences in the explode outcome condition. T1DM showed higher BOLD activity in areas related to error monitoring and uncertainty, in particular the medial prefrontal cortex and ACC.
2) Last balloon series (end of trial and error learning)
During the last balloon series, participants were deciding after iterative decision-making. Results for T1DM participants compared to control participants showed increased activations in saliency network regions related to error monitoring, such as bilateral ACC, and inhibitory control, such as left IFG. T1DM participants showed decreased activation in left posterior cingulate cortex and superior parietal lobe (Supplementary Table 2).
3) Cash out decision (to keep money and top risking)
Comparing T1DM versus control groups, cash out decision led to higher activity in frontal regions associated with decision-making (BA9, BA10, BA46), and saliency network regions such as insular cortex (bilateral insula). Participants from the T1DM group also showed higher activity than the control group in the hippocampus. Lower patient related activity was found in inhibitory control regions (right IFG BA45) inferior parietal lobe BA40 and posterior cingulate cortex.
4) Explode outcome (yielding full loss of reward)
Upon full loss outcomes, patients showed increased activation as compared to controls in frontal areas related to decision-making and error monitoring, namely the middle frontal gyrus (BA9, BA10) and ACC (BA24, BA32) regions within the saliency network.
Temporal trajectories of metabolic control are associated with activation in limbic and striatal circuits
We found positive correlations (meaning impairment) with trajectories metabolic control (as measured using the slope of changes of HbA1c over time) with similar areas as observed in above mentioned group contrasts for the first and last balloon periods: cingulate cortex (BA32, BA25) and in particular its subgenual limbic components and striatal regions (caudate). In the first balloon period we observed prominent correlated brain activation in frontal saliency regions involved in conflict monitoring such as the ACC. In the last balloon similar correlation patterns were observed in striatal and reward related regions (Caudate, putamen, NAcc) but now the insula, as part of the saliency network showed more significantly correlated activation. For negative rewards (explosion), negative correlations (related to better metabolic control) were found with posterior cingulate cortex (BA31, BA23), posterior parietal regions (BA7, BA39, BA40) and superior temporal lobe (BA22).
A very important sum up is that patterns related to successful trajectories metabolic control in patients and activation in healthy participants seem to be very much alike (matching parietal and posterior cingulate regions) in contrast with impaired metabolic control patients (distinct in frontal and anterior regions) (Fig. 3 and Supplementary Table 3).

A functional magnetic resonance imaging whole brain correlation analysis between glycosylated hemoglobin (HbA1c) values and blood oxygen level dependent (BOLD) activity, in type 1 diabetes mellitus patients. (A) Whole brain correlation analysis between BOLD activity during the first balloon condition and HbA1c (higher HbA1c trajectory values featuring a poorer metabolic control). A positive value (red) for the correlation, means that poorer the metabolic control (higher the HbA1c), higher the BOLD activity. This was found in prefrontal regions and in particular subgenual anterior cingulate cortex (ACC, 32, 25) and dorsal and ventral striatal regions. (B) Whole brain correlation analysis between BOLD activity during last balloon condition and HbA1c (higher HbA1c trajectories meaning a poorer metabolic control). A positive value (red) for the correlation, means that poorer the metabolic control (higher the HbA1c), higher the BOLD activity. This was found again in saliency regions (ACC, 32, 25) as well as in subcortical reward related regions (caudate, putamen, nucleus accumbens) and the Insula, whereas negative correlations were found mainly in posterior cingulate and parietal regions (BA31, BA23, BA7, BA39, BA40, but see temporal BA22).
Risk averse and risk seeking contrasts within T1DM
1) First balloon series and last balloon series
The contrast between risk averse and risk seeking during the first balloon and also for the last balloon series yielded larger activations for risk-taking (more adaptive decision) patients in parietal regions, suggesting, in line with above mentioned findings that higher activity in these regions is associated with better control (Fig. 4).

Group comparisons for patients with risk averse (less adaptive) versus risk-taking (learners by trial and error) profiles for the first balloon, last balloon, cash out decision and explode outcome conditions. (A) First balloon condition. Differences dominated in posterior parietal regions. (B) Last balloon condition. A similar pattern was observed (with the more adaptive patient mode being similar to the one observed in controls). (C) Cash out condition. Risk averse type 1 diabetes mellitus (T1DM) patients showed higher blood oxygen level dependent (BOLD) activity than risk seeking T1DM patients in insula, left BA44 and BA9. Conversely, risk seeking T1DM patients showed higher BOLD activity in right caudate, left anterior cingulate cortex (BA24, BA32), prefrontal cortex (BA10) and inferior frontal gyrus (BA44). (D) Explode outcome series. Risk averse T1DM patients showed higher BOLD activity than risk seeking T1DM patients in anterior cingulate cortex (BA32), inferior frontal gyrus (BA44), prefrontal cortex (BA9). Conversely, risk seeking T1DM patients showed higher BOLD activity than risk averse T1DM patients in parietal and posterior cingulate regions.
2) Cash outcome decision risk
Averse patients showed larger activations in insula, whereas risk seeking revealed activations in regions such as anterior cingulate gyrus and caudate, suggesting shifts of activity within the saliency network. Both groups shared activations in frontal inhibitory regions as middle (BA9, BA10) and IFG (BA44).
3) Explode outcome
ACC, middle and IFG regions involved in inhibitory control activated into a larger extent for risk averse T1DM patients in contrast with risk seeking profile, suggesting a distinct activation pattern in regions involved in cognitive and inhibitory control. Risk seeking T1DM patients showed higher blood oxygen level dependent (BOLD) activity than risk averse T1DM patients in parietal and posterior cingulate regions, further suggesting an important role for these regions (Supplementary Table 4).
DISCUSSION
We found triple converging indication that dopaminergic reward and saliency (interoceptive and error monitoring) circuits show a tight link with impaired metabolic trajectories and cognitive impulsivity in T1DM. Our findings suggest that distinct neuroimaging endophenotypes can be related to the success of metabolic control. These findings also have strong implications for programs to improve patient adherence, directly addressing risk-taking profiles. Accordingly, we found that T1DM, a chronic life-long disease, is associated with a modified neural risk processing, hindering adaptive impulse control behavior. We focus on four aspects of decision-making: (1) before trial and error learning; (2) after iterative decision-making; and at (3) cash out decisions (decision to take reward and not proceed); and (4) aversive outcomes, as balloon “explosions” (negative reward because no decision to cash out was made). We were particularly interested in dynamic profiles of HbA1c values over time and their relation with risk-taking brain networks. Accordingly, progressive variations from worse to the best control metabolic control (positive vs. negative slope, respectively) were related to brain activity patterns. These associations replicate the same pattern of group differences between T1DM and risk averse versus risk-taking (trial and error learning) profiles.
Group differences before and after iterative decision-making
We found that, in first balloon, patients activated into a larger extent the motivational system, which includes the NAcc and the amygdala [29,30]. Activity of the mesolimbic dopaminergic system that includes the ventral tegmental area which projects to NAcc and innervates as well the amygdala, is related to reactivity to emotional information and anticipation of reward [31]. Additionally, the IFG and insula are related to response inhibition and saliency processing for action selection, respectively. Patients preferred low rewards/losses which is distinct from studies with pathological gamblers [32].
Patients with worsening metabolic control showed increased activity in limbic, reward, and saliency regions. Importantly, they presented a distinct pattern of activations from patients with successful metabolic control who showed a pattern close to healthy participants brain activations, with dominant posterior cingulate and parietal recruitment, contrasting with patients with impaired metabolic control.
In general, our findings support an overactivation of brain regions related to motivation/reward and impulse control in T1DM patients. Under conditions of complete uncertainty and ambiguity and even after iterative decision-making, the pattern of choice did not change. Looking at behavioral data, patients prefer to cash out decisions related to avoidance or removal of aversive stimuli. Similar findings were found in obsessive compulsive disorder patients performing a computerized BART task [33]. Based on these data, the following interpretation can be put forward. (1) Patients miss the chance to get more information, from trial and error learning. This is important in the clinical context because in chronic disorders such operant way of learning new habits is very important. Patients respond aversively to negative outcomes (insula/ACC activation in saliency circuits) which promote anxiety (amygdala). (2) They are characterized by strong rigidity in action selection. (3) The trade-off between magnitude of a potential reward and probability of a negative outcome will eventually trigger inhibitory control processing and result in restricted behavioral patterns, precluding operant learning. (4) It is possible that abnormal frontal activation patterns may also explain maintenance of a rigid emotional state [34], thus preventing behavioral changes.
Appraisal and aversive outcomes
Risk averse and risk seeking performance in BART offered useful measures to understand extreme behavioral profiles. Patients with risk averse profile were exposed to more tension between reward seeking and loss aversion. This may explain the observation of dorsal striatum (caudate) and error monitoring circuit activations (ACC within the salience network) for cash out decisions (larger rewards) as well as inhibitory control and executive regions (BA44, BA 9/10). Conversely, risk-taking T1DM patients with adaptive exploratory behavior showed higher brain activity than risk averse T1DM patients in parietal and posterior cingulate regions, with we also show to be relevant to metabolic control and decision in healthy participants.
As a limitation of this study, with a larger sample subgroup stratification of risk could be better optimized. Second, we considered cash out decision and “explosion” outcomes in a general way. However, a cash out decision with lower reward brings a different tension from larger rewards. In the same way, a balloon explosion with few pumps (loosing less money) has a different impact than lose a huge amount of money. However, such fine grained analyses would require larger sample sizes.
Results of this research may offer insights to future directions concerning adaptive decision-making and impulse control in chronic life-long diseases such as diabetes. In this condition acquisition of adaptive patterns of behavior is important. Achievement of metabolic control is strongly dependent on adequate trial and error learning, and adequate impulse control in healthy habit acquisition. It will be helpful to discriminate if the biological status is a mediator or instead a consequence of the neural mechanisms that inhibit learning of appropriate behavioral responses, which is not possible to infer with correlative approaches. However previous work [1] identified a biological partition of metabolic control using K-means and clustering analysis, and the current work corroborates the notion that this partition can also be mapped from the neural point of view. The similarity with findings within anxiety spectrum disorders, as obsessive compulsive disorders [35] is quite interesting. It is likely that the biological worsening over time has an impact on cognitive flexibility that may explain suboptimal decision-making, as continuous oscillations of HbA1c have been also related to cognitive impairments. Interestingly, the systematic attention to disease control, also brings these patients closer to the typical pattern observed in post-traumatic stress disorder in which neuroimaging studies report excessive saliency processing, hyperactivity of anterior insula and dorsal anterior cingulate cortex and decreased top-down cognitive control involving fear and negative affect [36].
In conclusion, adaptive decision-making mechanisms related to cognitive impulsivity are affected in T1DM and are associated with the temporal trajectories of HbA1c. The neurobiological framework provided here suggests a link between reward, limbic and saliency networks in explaining biological worsening in patients with impaired metabolic control over time. Conversely, a posterior cingulate and parietal association to positive metabolic control was found. These results, which corroborate a matched biological partition of metabolic control trajectories. The observed behavioral and neuroimaging endophenotypes which could be related to the success of metabolic control have direct implications for programs to improve patient adherence. By directly addressing risk-taking profiles, trial and error learning and control of impulsivity in T1DM it is possible to design better patient education and monitoring programs.
SUPPLEMENTARY MATERIALS
Supplementary materials related to this article can be found online at https://doi.org/10.4093/dmj.2021.0307.
BART experiment: rational and description
Behavioral results on BART task (n=25), for risk averse and risk seeking performances within T1DM participants
Differences in brain activation for T1DM and controls between group analysis
Differences in brain activation within T1DM according to variations of metabolic control: correlation analysis
Differences in brain activation for risk averse and risk seeking patients between group analysis
Flowchart of functional magnetic resonance imaging predictors and study sample. T1DM, type 1 diabetes mellitus.
Performance at Balloon Analogue Risk Task (BART) on first balloon and last balloon series for type 1 diabetes mellitus (T1DM) and healthy participants, comparing two moments of experimental task: prior and after iterative decision making. T1DM group mean performance on first balloon was not significantly different from last balloon (unchangeable performance). Healthy participants do learn after iterative decision making) (P<0.05) and do become better than T1DM participants (P<0.05). CI, confidence interval.
Notes
CONFLICTS OF INTEREST
No potential conflict of interest relevant to this article was reported.
AUTHOR CONTRIBUTIONS
Conception or design: H.J., I.C.D., S.P., A.P.R., M.C.B.
Acquisition, analysis, or interpretation of data: H.J., I.C.D., S.P., A.P.R., M.C.B.
Drafting the work or revising: H.J., I.C.D., S.P., A.P.R., M.C.B.
Final approval of the manuscript: M.C.B.
FUNDING
This work was supported by Fundação para a Ciência e Tecnologia—FCT (UID/4950/2020, DSAIPA/DS/0041/2020, PCIF/SSO/0082/2018, PTDC/PSI-GER/30852/2017, PTDC/PSI-GER/1326/2020, INFARMED Research Fund for Health (FIS-FIS-2015-01 DIA - DiaMarkData), and the European Foundation for the Study of Diabetes (EFSD) 2019—Innovative Measurement of Diabetes Outcomes 2019.
Acknowledgements
We would like thank all participants for their contribution to our study.