- Current
- Browse
- Collections
-
For contributors
- For Authors
- Instructions to authors
- Article processing charge
- e-submission
- For Reviewers
- Instructions for reviewers
- How to become a reviewer
- Best reviewers
- For Readers
- Readership
- Subscription
- Permission guidelines
- About
- Editorial policy
Articles
- Page Path
- HOME > Diabetes Metab J > Volume 44(1); 2020 > Article
-
Original ArticleBasic Research Fibroblast Growth Factor 21 Attenuates Diabetes-Induced Renal Fibrosis by Negatively Regulating TGF-β-p53-Smad2/3-Mediated Epithelial-to-Mesenchymal Transition via Activation of AKT
-
Sundong Lin1,2,3
 , Lechu Yu1, Yongqing Ni1, Lulu He1,2,3, Xiaolu Weng1,2, Xuemian Lu1, Chi Zhang1,2,3
, Lechu Yu1, Yongqing Ni1, Lulu He1,2,3, Xiaolu Weng1,2, Xuemian Lu1, Chi Zhang1,2,3 -
Diabetes & Metabolism Journal 2020;44(1):158-172.
DOI: https://doi.org/10.4093/dmj.2018.0235
Published online: October 28, 2019
1Ruian Center of Chinese-American Research Institute for Diabetic Complications, The Third Affiliated Hospital of Wenzhou Medical University, Wenzhou, China.
2Chinese-American Research Institute for Diabetic Complications, Wenzhou Medical University, Wenzhou, China.
3School of Pharmaceutical Science, Wenzhou Medical University, Wenzhou, China.
- Corresponding author: Chi Zhang. Chinese-American Research Institute for Diabetic Complications, Wenzhou Medical University, Wenzhou 325035, China. zhangchi515@126.com
- Corresponding author: Xuemian Lu. Ruian Center of Chinese-American Research Institute for Diabetic Complications, Wenzhou Medical University, Wenzhou 325200, China. lu89118@medmail.com.cn
- *Sundong Lin and Lechu Yu contributed equally to this study as first authors.
Copyright © 2020 Korean Diabetes Association
This is an Open Access article distributed under the terms of the Creative Commons Attribution Non-Commercial License (http://creativecommons.org/licenses/by-nc/4.0/) which permits unrestricted non-commercial use, distribution, and reproduction in any medium, provided the original work is properly cited.
ABSTRACT
-
Background
- Epithelial-to-mesenchymal transition (EMT) is required for renal fibrosis, which is a characteristic of diabetic nephropathy (DN). Our previous study demonstrated that fibroblast growth factor 21 (FGF21) prevented DN associated with the suppressing renal connective tissue growth factor expression, a key marker of renal fibrosis. Therefore, the effects of FGF21 on renal fibrosis in a DN mouse model and the underlying mechanisms were investigated in this study.
-
Methods
- Type 1 diabetes mellitus was induced in C57BL/6J mice by intraperitoneal injections of multiple low doses of streptozotocin. Then, diabetic and non-diabetic mice were treated with or without FGF21 in the presence of pifithrin-α (p53 inhibitor) or 10-[4′-(N,N-Diethylamino)butyl]-2-chlorophenoxazine hydrochloride (10-DEBC) hydrochloride (Akt inhibitor) for 4 months.
-
Results
- DN was diagnosed by renal dysfunction, hypertrophy, tubulointerstitial lesions, and glomerulosclerosis associated with severe fibrosis, all of which were prevented by FGF21. FGF21 also suppressed the diabetes-induced renal EMT in DN mice by negatively regulating transforming growth factor beta (TGF-β)-induced nuclear translocation of Smad2/3, which is required for the transcription of multiple fibrotic genes. The mechanistic studies showed that FGF21 attenuated nuclear translocation of Smad2/3 by inhibiting renal activity of its conjugated protein p53, which carries Smad2/3 into the nucleus. Moreover pifithrin-α inhibited the FGF21-induced preventive effects on the renal EMT and subsequent renal fibrosis in DN mice. In addition, 10-DEBC also blocked FGF21-induced inhibition of renal p53 activity by phosphorylation of mouse double minute-2 homolog (MDM2).
-
Conclusion
- FGF21 prevents renal fibrosis via negative regulation of the TGF-β/Smad2/3-mediated EMT process by activation of the Akt/MDM2/p53 signaling pathway.
- Diabetic nephropathy (DN) is one of the most severe complications of diabetes [1]. Approximately 40% of patients with DN develop end-stage renal disease (ESRD) [2]. Progression of DN is characterized by gradual renal fibrosis, which is attributed to the excessive accumulation of the extracellular matrix (ECM) manifested by the overexpression of matrix proteins, including connective tissue growth factor (CTGF), fibronectin 1 (FN1), and collagen [3]. In diabetes, ECM deposition in the glomerulus and tubular-interstitial compartment is mainly responsible for tubulointerstitial fibrosis and glomerulosclerosis [4].
- Strong evidence has demonstrated that ECM deposition is primarily induced by the epithelial-to-mesenchymal transition (EMT), a process in which epithelial cells transdifferentiate into motile mesenchymal cells [5]. Epithelial cells play important roles in maintaining normal renal structure and function. However, during the EMT, epithelial cells gradually lose apical-basal polarity and intracellular adhesions, along with acquisition of the mesenchymal cell phenotype such as migratory and invasive abilities. This process is also characterized by the decreased expression of specific epithelial proteins, including E-cadherin and zonula occludens-1 (ZO-1), followed by the increased expression of mesenchymal-specific proteins, including α-smooth muscle actin (α-SMA), vimentin, and laminin [6]. Moreover, accumulating studies have confirmed that the inhibition of the EMT in the kidney significantly suppresses the progression of renal fibrosis and subsequent DN [5].
- Mechanistic studies have indicated that the transforming growth factor beta (TGF-β)-p53-Smad2/3 pathway plays a critical role in mediating the EMT and renal fibrosis [78]. Sustained high-glucose levels induce the renal expression of TGF-β, which activates Smad 2/3 by phosphorylation. Then, phosphorylated Smad2/3 translocates into the nuclei from the cytosol and functions as transcription factors that induce the expression of multiple fibrotic genes, including CTGF, TGF-β, and plasminogen activator inhibitor-1 (PAI-1) [79]. During the above-mentioned process, p53 is required for Smad2/3 nuclear translocation. Activated p53 assembles and binds to TGF-β-activated Smad2/3 and forms a complex that is able to translocate across the nuclear membrane [79]. Therefore, suppression of the EMT by inhibiting the p53-mediated Smad2/3 nuclear translocation may be an effective strategy for preventing diabetes-induced renal fibrosis and subsequent DN.
- Fibroblast growth factor 21 (FGF21) is an important metabolic regulator that induces preventive effects against diabetes and its complications [10]. Our previous study indicated that the administration of FGF21 attenuated type 1 or type 2 diabetes mellitus-induced early-stage renal apoptosis and late-stage DN [11]. Although we found that FGF21 significantly suppressed CTGF expression in the diabetic kidney, CTGF is just one of multiple markers of renal fibrosis at the molecular level. It is not sufficient to conclude that FGF21 induces anti-fibrotic effects in the kidney of DN mice. If so, whether its anti-fibrotic effects are attributed to the suppression of the EMT is still unclear. Additionally, strong evidence has shown that FGF21 is a negative regulator of p53 [12]. Wang et al. [13] reported that FGF21 represses cerebrovascular aging by improving mitochondrial biogenesis and inhibiting the p53 signaling pathway. Another study demonstrated that FGF21 suppresses cisplatin-induced kidney tubular cell injury by suppressing p53 induction and activity [12]. In contrast, a decrease of endogenous FGF21 in the liver is associated with aberrant p53 and TGF-β/Smad signaling during hepatocellular carcinoma development [14]. Moreover, increasing evidence has demonstrated that p53 activity is negatively regulated by the activation of Akt, which is the classic downstream target enzyme of FGF21 [15].
- In this study, we mainly focused on test the hypothesized that FGF21 prevents renal fibrosis and subsequent DN by the activation of Akt-mediated suppression of the TGF-β-p53-Smad2/3-induced renal EMT.
INTRODUCTION
- Ethics statement
- The experimental protocol was approved by the Committee on the Ethics of Animal Experiments of Wenzhou Medical University (IRB number: SYXK2015-0009, Zhejiang, China). The animals were killed under anesthesia induced by intraperitoneal injection of 1.2% 2,2,2-tribromoethanol (avertin) (Sigma-Aldrich, St. Louis, MO, USA) at a dose of 20 µL/g body weight, and all efforts were made to minimize their suffering.
- Animal model and treatment
- Type 1 diabetes mellitus was induced in wild type (WT) and FGF21 knockout (FGF21-KO) mice (8 weeks old, male) with the background of C57BL/6J by intraperitoneal injection of streptozotocin at a dose of 50 mg/kg/day for 5 consecutive days, while age-matched control mice received injections of the same volume of sodium citrate buffer [16]. After hyperglycemia was stably diagnosed, diabetic mice (WT) were treated with or without FGF21 (produced in our laboratory, 100 µg/kg/day) in the presence of pifithrin-α (PF-α) (intraperitoneally administered at 1.1 mg/kg five times a week; Sigma-Aldrich) [1011] or 10-[4′-(N,N-Diethylamino)butyl]-2-chlorophenoxazine hydrochloride (10-DEBC) (intraperitoneally administered at 1 mg/kg seven times a week; Sigma-Aldrich) for 4 months. Same volume of phosphate-buffered saline was used as vehicle. The grouping is as following: control (Con) group (n=8), FGF21 group (n=8), DN group (n=8), DN/FGF21 group (n=8), DN/PF group (n=8), DN/DEBC group (n=8), DN/FGF21/PF group (n=8), and DN/FGF21/DEBC group (n=8). The technical route was shown in Supplementary Fig. 1.
- Measurements of renal function
- Four months after FGF21 and/or PF-α treatment, 24-hour urine collection was done from mice housed in metabolic cages, and the sera were collected after the mice were killed. Blood urea nitrogen (BioAssay Systems, Hayward, CA, USA), urine protein (BioAssay Systems), urine microalbumin (Bethyl Laboratories, Montgomery, TX, USA), and serum creatinine (BioAssay Systems) were measured according to the manufacturers' instructions. The urinary albumin-to-creatinine ratio was expressed as urine albumin/urine creatinine (µg/mg).
- Histological staining
- The renal tissue of mice was fixed in 10% formalin at room temperature for 48 hours. After dehydration, the tissue blocks were embedded in paraffin and cut into 4-mm-thick blocks, followed by hematoxylin and eosin (H&E) staining for general morphological examination, periodic acid–Schiff (PAS) staining to evaluate glomerulosclerosis, and Sirius red staining and Masson trichrome staining to evaluate collagen accumulation, which represent fibrosis [1718].
- Nuclei isolation
- The nuclei of renal cells were isolated using the Nuclei Isolation Kit (NUC-201; Sigma-Aldrich) as previously described [19]. Briefly, 50 mg renal tissue was homogenized for 50 seconds in 300 mL cold lysis buffer containing 1 mL dithiothreitol and 0.1% Triton X-100. Then, 600 mL of cold 1.8 M Cushion Solution was added to the lysis solution. The mixture was transferred to a new tube pre-loaded with 300 mL of 1.8 M Sucrose Cushion Solution followed by centrifugation at 30,000 ×g for 45 minutes. The supernatant containing the cytoplasmic component was saved for later analysis. Nuclei were visible as a thin pellet at the bottom of the tube.
- RNA isolation and quantitative polymerase chain reaction
- Total RNA was isolated from kidney tissue using TRIzol reagent (Invitrogen, Carlsbad, CA, USA). The concentration of total mRNA in each sample was quantified with the Nanodrop 2000 Spectrometer (Thermo Fisher Scientific, San Jose, CA, USA). The mRNA samples were reverse transcribed into cDNA using the High-Capacity cDNA Reverse Transcription Kit (PE Applied Biosystems, Foster City, CA, USA). Quantitative polymerase chain reaction was conducted in triplicate using the SYBR GREEN PCR master mix (Invitrogen) on the Stratagene MX3000p Thermocycler (Agilent StrataGene, Santa Clara, CA, USA). The amount of mRNA was calculated by the comparative computed tomography (CT) method, which depends on the ratio of the amount of target genes to the reference gene β-actin.
- Western blot analysis
- Renal tissues were homogenized in lysis buffer (Santa Cruz Biotechnology, Santa Cruz, CA, USA), followed by centrifugation at 12,000 ×g at 4℃ to collect the supernatant. Equal amounts of protein from each sample were separated on 10% sodium dodecyl sulfate polyacrylamide gel electrophoresis and transferred to nitrocellulose membranes. After blocking in non-fat milk for 1 hour at room temperature, membranes were incubated overnight at 4℃ with the following primary antibodies: CTGF (1:1,000), FN1 (1:1,000), PAI-1 (1:1,000), collagen I (Col I, 1:1,000), collagen II (Col II, 1:1,000), collagen IV (Col IV, 1:1,000), E-cadherin (1:1,000), phosphorylated cadherin (1:1,000), nephrin (1:1,000), ZO-1 (1:1,000), α-SMA (1:1,000), vimentin (1:1,000), laminin (1:1,000), TGF-β (1:1,000), p-Smad2 (1:1,000), total Smad2 (t-Smad2, 1:1,000), p-Smad3 (1:1,000), t-Smad3 (1:1,000), p-Smad7 (1:1,000), t-Smad7 (1:1,000), snail (1:1,000), twist (1:1,000), slug (1:1,000), phosphorylated mouse double minute-2 homolog (p-MDM2, 1:1,000), total MDM2 (1:1,000), lamin B (1:1,000), and β-actin (1:1,000) (all purchased from Abcam, Cambridge, MA, USA), p-p53 (1:1,000) and t-P53 (1:1,000) were purchased from Cell Signaling Technology (Danvers, MA, USA). After three washes in Tris-buffered saline containing 0.05% Tween 20, the membranes were incubated with horseradish peroxidase-conjugated secondary antibodies for 1 hour at room temperature. Antigen-antibody complexes were visualized using an enhanced chemiluminescence kit (Amersham, Piscataway, NJ, USA), and the intensity of the protein bands was quantified using Quantity One software (Bio-Rad, Hercules, CA, USA).
- Immunoprecipitation
- Immunoprecipitation (IP) was performed with kidney tissues (30 mg from each mouse) lysed in IP buffer using the Pierce Co-IP Kit (Pierce Biotechnology Ltd., Rockford, IL, USA) as previously described [2021]. Immunocomplexes were determined using specific antibodies as indicated.
- Statistical analysis
- Data were collected from eight mice per group, and are presented as the mean±standard deviation. One-way analysis of variance was used to determine general differences, followed by the post hoc Tukey's test to determine the difference between groups. Origin 7.5 software (OriginLab Corp., Northampton, MA, USA) was used for laboratory data analysis and graphing. Statistical significance was considered P<0.05.
METHODS
- FGF21 suppresses renal hypertrophy and maintains renal function
- Renal hypertrophy and dysfunction are the key symptoms of DN, which were examined to evaluate the preventive effects of FGF21 on DN in this study. We found that the ratio of kidney weight to tibia length significantly increased, indicating renal hypertrophy in DN mice (Supplementary Table 1). Meanwhile, renal function was also impaired in 4-month-old DN mice, as characterized by an increase of urine volume, urine protein content, blood creatinine level, urine microalbumin level, blood urine nitrogen level, and the ratio of urinary albumin/urinary creatinine (Supplementary Table 1). However, the parameters representing renal hypertrophy and renal dysfunction were strongly suppressed by administration of FGF21 (Supplementary Table 1), indicating that FGF21 treatment induces preventive effects on DN. Additionally, FGF21 treatment slightly decreased blood glucose level and glycosylated hemoglobin in type 1 diabetes mellitus mice (Supplementary Table 1).
- FGF21 prevents renal morphological changes, glomerulosclerosis, and fibrosis in the diabetic kidney
- Generally, diabetes-induced renal dysfunction is associated with renal pathological changes. H&E staining (Supplementary Fig. 2A) and PAS staining (Supplementary Fig. 2B and E) showed that pathological changes were obvious in the diabetic kidney and included tubular atrophy, tubulointerstitial lesions, glomerular expansion, glomerulosclerosis, mesangial cell proliferation, mesangial matrix expansion, and capillary collapse, all of which were suppressed by FGF21 treatment for 4 months (Supplementary Fig. 2A, B, and E). In addition, Sirius red staining (Supplementary Fig. 2C and F) and Masson staining (Supplementary Fig. 2D and G) showed clear collagen accumulation in both the glomeruli and tubule, representing glomerular and tubulointerstitial fibrosis, respectively. In contrast, diabetes-induced renal fibrosis was prevented by 4 months of FGF21 treatment (Supplementary Fig. 2 C, D, F, and G).
- FGF21 prevents diabetic renal fibrosis associated with suppression of renal ECM accumulation
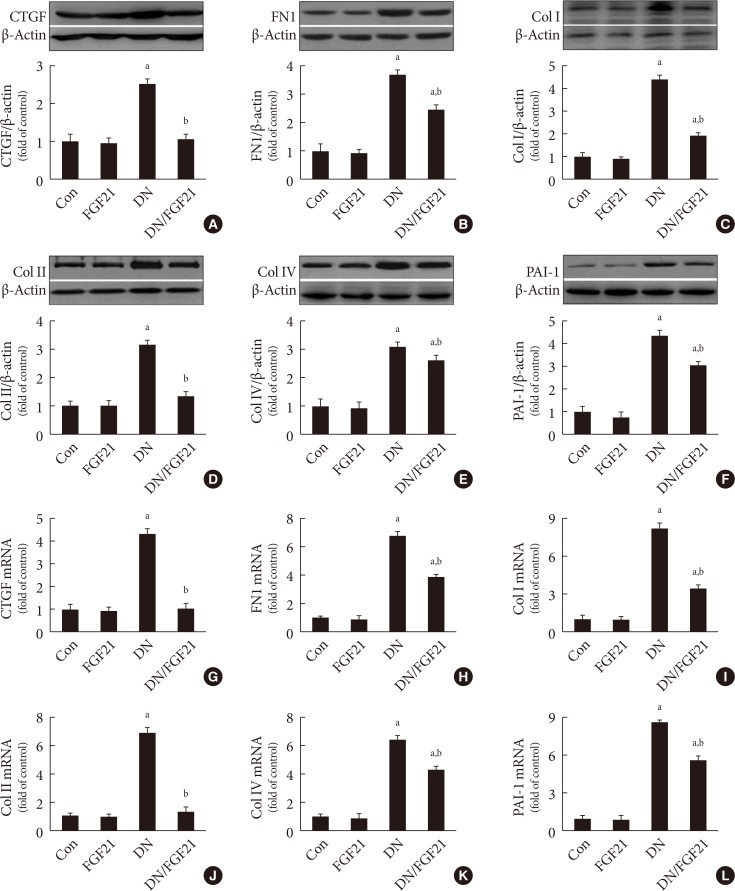
- Since the ECM is the predominant contributor to diabetic renal fibrosis, the markers represented ECM were examined in this study. The expression of CTGF, a strong inducer of ECM synthesis, was significantly increased in the diabetic kidney (Fig. 1A). Meanwhile, the expression of multiple classic matrix proteins in the diabetic kidneys, including FN1 (Fig. 1B), collagen of types I (Fig. 1C), II (Fig. 1D), and IV (Fig. 1E), were significantly increased. Additionally, the expression of PAI-1, considered both an inflammatory and fibrotic marker, was strongly increased (Fig. 1F). Furthermore, the quantitative polymerase chain reaction assay showed that the mRNA levels of these ECM markers above were also increased in the diabetic kidney (Fig. 1G–L), indicating that the upregulated ECM markers were derived from the kidney rather than transferred into the kidney in an endocrine manner. However, FGF21 treatment for 4 months notably inhibited the increased ECM mRNA and protein expression in the diabetic kidney (Fig. 1G–L), indicating that FGF21-induced anti-fibrotic effect might be attributed to their suppression of the ECM in the diabetic kidney.
- FGF21 prevents the EMT in the diabetic kidney
- Next, we examined the effects of FGF21 on the EMT, which is the predominant inducer of ECM accumulation and fibrosis in the diabetic kidney. To this end, the protein expression levels of EMT biomarkers in the renal cortex were detected by Western blotting. The results showed that the EMT was significantly enhanced in the diabetic kidney, as characterized by a decrease in the expression of epithelial cells markers, including E-cadherin (Supplementary Fig. 3A and B) and ZO-1 (Supplementary Fig. 3A and C), and an increase in the expression of mesenchymal cells markers, including α-SMA (Supplementary Fig. 3A and D), vimentin (Supplementary Fig. 3A and E), and laminin (Supplementary Fig. 3A and F). However, the EMT process above was prevented by FGF21 treatment (Supplementary Fig. 3A–F). Next, we found that the FGF21-induced preventive effects on EMT in the diabetic kidney were associated with the inhibition of snail (Supplementary Fig. 3A and G), twist (Supplementary Fig. 3A and H), and slug (Supplementary Fig. 3A and I), which are negative transcriptional regulators of E-cadherin. Generally, studies related to EMT focus on tubular cells; however, in this study, we found that FGF21 treatment also preserved the renal expression of P-cadherin (Supplementary Fig. 3A and J), which is derived from podocytes in the diabetic kidney. Therefore, we also detected the expression of nephrin, a biomarker of podocytes that are classified as epithelial cells in the glomeruli. Nephrin expression was strongly inhibited in the diabetic kidney (Supplementary Fig. 3A and K), indicating that the EMT occurs in both the tubules and glomeruli of the diabetic kidney. Although under healthy condition, FGF21 treatment had no impact on EMT, which were notably suppressed under diabetic condition (Supplementary Fig. 3).
- FGF21 negatively regulates Smad2/3 nuclear translocation and renal fibrosis by inhibiting p53 activity
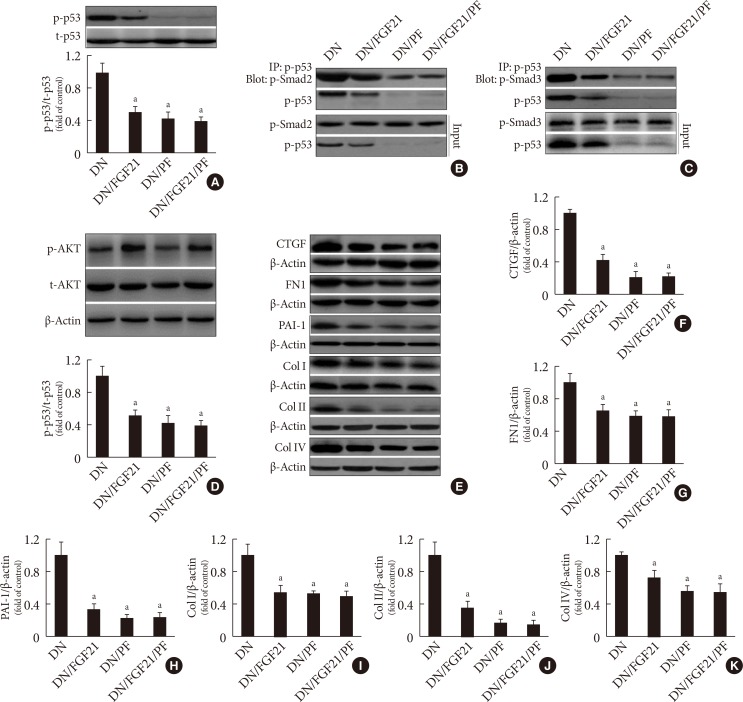
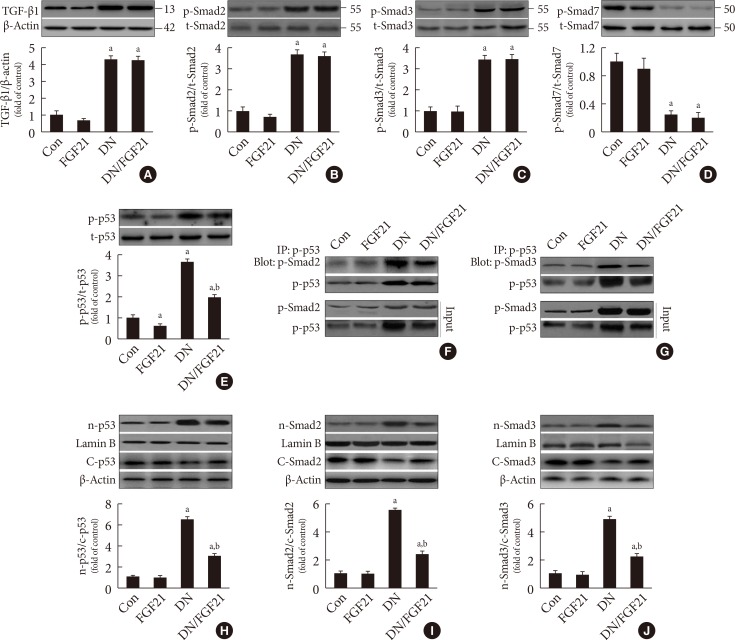
- In this study, we found that diabetes strongly increased the expression of PAI-1, a classic marker of TGF-β signaling, at both the mRNA and protein levels. Importantly, TGF-β/Smad2/3 signaling is the predominant regulatory pathway of the EMT in the kidney. Western blot analyses showed that TGF-β expression was significantly upregulated in diabetic conditions (Fig. 2A), followed by activity (phosphorylation) enhancement of Smad2 and Smad3 in the diabetic kidney (Fig. 2B and C). Opposite changing pattern of the phosphorylation renal Smad7, negative regulator of Smad2/3 (Fig. 2D) was observed in diabetic mice. Unexpected, FGF21 treatment had no impact on the expression of TGF-β (Fig. 2A) and phosphorylation of Smad2/3/7 in the diabetic kidney (Fig. 2B–D), indicating that FGF21 does not affect TGF-β/Smad2/3 signaling transduction. Strong evidence has demonstrated that p53 is a key kinase that regulates TGF-β-Smad2/3 by binding with Smad2/3. This study showed that diabetes induced p53 phosphorylation in the diabetic kidney (Fig. 2E). Meanwhile, the binding amount of phosphorylated p53 to phosphorylated Smad2/3 significantly increased in the diabetic kidney (Fig. 2F and G). However, FGF21 treatment for 4 months not only suppressed p53 phosphorylation (Fig. 2E), but also attenuated the binding amount of p-p53 and p-Smad2/3 (Fig. 2F and G). Next, we evaluated the nuclear translocation of the p53/Smad2/3 complex by detecting the ratio of nuclear content to cytosol content of both p53 and Smad2/3, which were significantly increased in the diabetic kidney (Fig. 2H–J). However, translocation of the complex was inhibited in the presence of FGF21 (Fig. 2H–J). In order to further confirmed the negative regulating effect of FGF21 on renal p53 activity and its nuclear translocation with Smad2/3, reverse experiments was performed in FGF21-KO mice compared with WT mice. The results showed that compared with WT diabetic mice FGF21 deletion further enhanced diabetes-induced renal p53 phosphorylation (Supplementary Fig. 4A) and its nuclear translocation (Supplementary Fig. 4B). Similarly, enhanced nuclear translocation of Smad2/3 were also observed in the diabetic kidney of FGF21-KO mice (Supplementary Fig. 4C and D), indicating that endogenous FGF21 negatively regulated p53-Smad pathway in the diabetic kidney.
- To determine the reason for the decreased interaction between p53 and Smad2/3 in the FGF21-treated diabetic kidney, PF-α (p53 specific inhibitor) was applied. The results showed that FGF21 or PF-α significantly suppressed p53 phosphorylation (Fig. 3A) and the protein-protein interaction between p53 and Smad2/3 (Fig. 3B and C). However, within treatment of PF-α, FGF21 failed to decrease p53 phosphorylation and the binding amount to Smad2/3 (Fig. 3), indicating that the FGF21-mediated decrease in the interaction between p53 and Smad2/3 was purely due to inhibition of the activity of p53 rather than suppressing the binding ability of p53 to Smad2/3. Additionally, we found inhibition of p53 had no effect on FGF21-induced renal AKT activity (Fig. 3D), indicating AKT is not the downstream target of p53. Moreover, diabetes-induced renal fibrosis and ECM accumulation were notably prevented at varying degrees by FGF21 or PF-α treatment, respectively, accompanied by the decreased expression of fibrotic markers and ECM markers, including CTGF (Fig. 3E and F), FN1 (Fig. 3E and G), PAI-1 (Fig. 3E and H), Col I (Fig. 3E and I), Col II (Fig. 3E and J), and Col IV (Fig. 3E and K). With PF-α treatment, FGF21 failed to further enhance the prevention of fibrosis and ECM accumulation in the diabetic kidney (Fig. 3E–K), indicating that FGF21 prevented diabetes-induced EMT in the kidney via inhibition of renal p53 activity.
- FGF21 prevents the renal EMT via inhibition of p53 activity in DN mice
- Since we found that FGF21 inhibited the TGF-β/Smad2/3 pathway by attenuating p53-mediated Smad2/3 nuclear translocation, we next examined the role of p53 in the FGF21-induced preventive effects of the EMT in the diabetic kidney. The results showed that diabetes-induced EMT in the kidney was notably prevented by FGF21 or PF-α treatment at varying degrees, with characteristics of increased expression of epithelial cell markers, including E-cadherin (Supplementary Fig. 5A and B), P-cadherin (Supplementary Fig. 5A and C), nephrin (Supplementary Fig. 5A and D), and ZO-1 (Supplementary Fig. 5A and E), and the decreased expression of α-SMA (Supplementary Fig. 5A and F), vimentin (Supplementary Fig. 5A and G), and laminin (Supplementary Fig. 5A and H). As expected, with PF-α treatment, FGF21 failed to further attenuate renal EMT in DN mice (Supplementary Fig. 5), indicating that FGF21 suppresses diabetes-induced EMT in the diabetic kidney via inhibition of renal p53 activity.
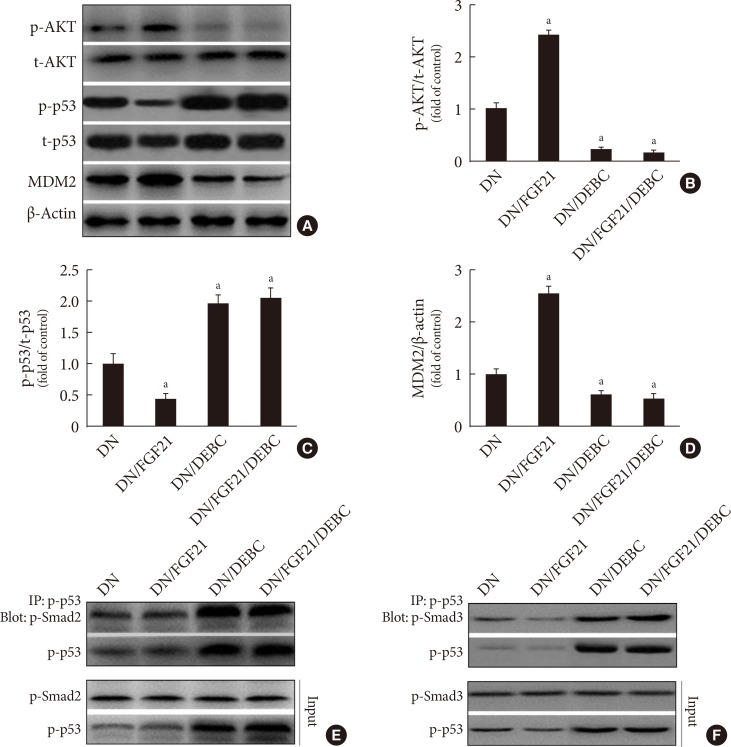
- AKT mediates FGF21-induced suppression of p53 activity and its binding to Smad2/3 by upregulating MDM2 expression
- To identify the role of Akt in FGF21-induced negative regulation of p53 activity in the diabetic kidney, a specific AKT inhibitor, 10-DEBC, was applied. The results showed that FGF21 enhanced, but DEBC suppressed AKT phosphorylation rather than its expression in the diabetic kidney (Fig. 4A and B), indicating that renal AKT inhibition model was successfully established. Additionally, FGF21 notably decreased, but 10-DEBC increased p53 phosphorylation in the diabetic kidney (Fig. 4A and C). However, the converse was observed with renal MDM2 in DN mice (Fig. 4A and D), suggesting that FGF21 suppresses renal p53 phosphorylation by increasing AKT-mediated MDM2 activation. Additionally, inhibition of Akt enhanced formation of the p53/Smad2/3 complex (Fig. 4E and F). Interestingly, FGF21 treatment failed to induce inhibitory effects on p53 phosphorylation and subsequent binding to Smad2/3 in the presence of 10-DEBC (Fig. 4), indicating that Akt is the key enzyme that negatively regulates p53/Smad2/3 signaling.
- FGF21 prevents diabetes-induced EMT and subsequent fibrosis in the kidney via activation of Akt
- The results showed that 10-DEBC treatment enhanced diabetes-induced renal EMT by further decreasing the expression of epithelial cells markers, including E-cadherin (Fig. 5A), P-cadherin (Fig. 5B), and ZO-1 (Fig. 5C), as well as further increasing the expression of mesenchymal cell markers, including α-SMA (Fig. 5D), vimentin (Fig. 5E), and laminin (Fig. 5F). Moreover, 10-DEBC treatment also enhanced diabetes-induced renal fibrosis, characterized by a further increase in the expression of multiple fibrotic and EMC markers, including CTGF (Fig. 5G), FN1 (Fig. 5H), PAI-1 (Fig. 5I), Col I (Fig. 5J), Col II (Fig. 5K), and Col IV (Fig. 5L). As expected, FGF21 was incapable of inducing preventive effects on renal EMT and fibrosis in the diabetic kidney in the presence of 10-DEBC (Fig. 5), indicating that Akt plays a key role in mediating FGF21-induced anti-fibrotic effects in the diabetic kidney.
RESULTS
- FGF21 is a member of FGF family, which induces beneficial effects in multiple organs in an endocrine manner [22]. Increasing clinical studies have demonstrated that circulating FGF21 levels are dramatically increased in patients with various kidney diseases, including chronic kidney disease (CKD) [2324], kidney transplantation [2526], ESRD [27], early-stage diabetic kidney disease [28], acute renal dysfunction [29], and long-term peritoneal dialysis [30], indicating that an increase in FGF21 levels above baseline is a stress response to induce renal protection. This hypothesis was confirmed by a subsequent study, which showed that the administration of FGF21 maintained renal function in mice with CKD [31]. Kim et al. [32] reported that FGF21 improved insulin resistance and ameliorated renal injury in spontaneous type 2 diabetes mellitus (db/db) mice. Our previous studies also indicated that FGF21 supplement prevents, but FGF21 deletion enhances DN progression [1117]. Importantly, we found that FGF21 supplement suppresses renal CTGF expression in the diabetic kidney [2733]. However, CTGF is only one marker of fibrosis; thus, the effects of FGF21 on renal CTGF expression are not enough to identify the anti-fibrotic effects of FGF21 in the kidney of DN mice, which were evaluated in this study.
- Four months after diabetes was diagnosed, the symptoms of DN were clearly observed. Meanwhile, fibrosis and it-induced glomerulosclerosis also obviously appeared in tubular and glomerular areas. The above findings indicate that fibrosis developed in both the tubules and glomerulus. Moreover, renal fibrosis was further confirmed by the increased expression of multiple fibrosis and ECM markers in the diabetic kidney. However, all of the above-mentioned symptoms were markedly prevented by the administration of FGF21, indicating that FGF21 induced anti-fibrotic effects in the kidney of DN mice.
- Although FGF21 is considered as metabolic regulator, in the present study only a slight glucose-lowering effect (approximately 10%) of FGF21 with the dose of 100 µg/kg was found in type 1 diabetes mellitus mice, which may not be the predominant contributor to the strong anti-fibrotic effect of FGF21 in the diabetic kidney indicating other mechanism must exist. Strong evidence has indicated that the EMT plays a significant role in renal fibrosis and ECM accumulation, which is a hallmark of DN [5]. The EMT can be divided into three different types according to the different biological circumstances in which they occur and the associated consequences [3435]. Among them, the type 2 EMT is implicated in wound healing, organ fibrosis, and tissue regeneration, and induces the generation of activated mesenchymal cells, which stimulate ECM accumulation ultimately leading to tissue damage [3637]. The EMT occurs in the diabetic kidney and is classified as type 2 EMT, which is considered the inducer of diabetic renal fibrosis [34].
- In this study, we also confirmed the enhanced EMT in the diabetic kidney with characteristics of decreased expression of epithelial cells markers and increased expressions of mesenchymal cells markers. Normally, the EMT in the kidney refers to tubular EMT [8], but the EMT in glomeruli also plays important roles in inducing podocyte loss and glomerular damage [3839], which was observed in this study. Importantly, the expression of all of the above-mentioned EMT markers was oppositely regulated by the administration of FGF21, indicating that FGF21 induced preventive effects on the EMT in both the tubular and glomerular areas of DN mice. Next, we focused on determining the underlying mechanisms of FGF21-induced suppression on the EMT in the diabetic kidney. EMT-induced renal fibrosis is predominantly mediated by the TGF-β/Smad2/3 pathway [4041]. In this study, we confirmed activated TGF-β/Smad2/3 in the diabetic kidney, characterized by increased renal TGF-β expression and phosphorylation of its downstream Smad2/3. Unexpectedly, FGF21 supplement inhibited TGF-β/Smad2/3-mediated EMT via the suppression of Smad2/3 nuclear translocation rather than inhibiting the expression or activity of TGF-β/Smad2/3. Importantly, we found that renal p53 is a key target of FGF21 that decreases activated Smad2/3 nuclear translocation. Increasing studies have reported that p53 plays an important role in regulating the TGF-β/Smad2/3 pathway [4243]. Activated p53 recognizes and binds to Smad2/3 phosphorylated by TGF-β to form a complex that is able to translocate into the nucleus and acts as a transcription factor of multiple fibrotic genes [4243]. Moreover, in vitro and in vivo studies have demonstrated that FGF21 can induce inhibitory effects on p53 activity [134445]. Similar effects were also identified in this study, in that FGF21 supplement suppressed renal p53 phosphorylation in the diabetic kidney, which contributed to less activated p53 binding to phosphorylated Smad2/3 to form the transcriptional complex. We also found that FGF21 supplement failed to reduce the amount and nuclear translocation of p53-Smad2/3 complex in the presence of PF-α, suggesting that suppression of p53 is required in FGF21-induced negative regulation of Smad2/3 nuclear translocation. Next, we mainly focused on determining whether FGF21-induced downregulation of Smad2/3 nuclear translocation via inhibition of p53 contributes to the suppression of EMT and subsequent renal fibrosis. We found that FGF21 supplement was incapable of inducing preventive effects on diabetes-induced renal EMT process and fibrosis in DN mice in the presence of PF-α, suggesting that FGF21 negatively regulated the EMT and subsequent fibrosis in the diabetic kidney by inhibition of the p53-mediated TGF-β/Smad2/3 pathway.
- Finally, we explored the underlying mechanisms of FGF21-induced negative regulation of renal p53 activity in the DN mice. Akt, an effector of phosphoinositide 3-kinase, is a serine/threonine protein kinase that regulates a variety of cellular functions and induces multiple beneficial effects in DN [4647]. AKT negatively regulates p53 activity by phosphorylating MDM2, a p53 negative regulator [48]. Then, activated MDM2 translocates into the nucleus and assembles phosphorylated p53. MDM2 carries p53 out of the nucleus for degradation [4950]. In this study, we found that FGF21 supplement increased the phosphorylation of renal MDM2. However, this effect was not observed in the presence of 10-DEBC, indicating that AKT is required for FGF21-induced activation of MDM2. Moreover, 10-DEBC also blocked FGF21-induced suppression of renal p53 and subsequent EMT and fibrosis in the diabetic kidney.
- In summary, we confirmed that FGF21 attenuates DN associated with the prevention of diabetes-induced renal ECM accumulation and fibrosis. Mechanistic studies indicated that FGF21 suppressed renal fibrosis in the diabetic kidney by negatively regulating TGF-β-p53-Smad2/3-mediated EMT via activation of Akt.
DISCUSSION
-
Acknowledgements
- This work was supported by grants from the Medical and Healthy Technological Grant of Zhejiang Province (Nos. 2015KYB236 to Chi Zhang and 2018KY769 to Lechu Yu), the National Science Foundation of China (No. 81670767 to Chi Zhang and 81700732 to Lechu Yu), and the Project for Selected Overseas Chinese supported by Zhejiang Technology Foundation to Chi Zhang. The funding institutions had no role in the study design, data collection and analysis, decision to publish, or preparation of the manuscript.
ACKNOWLEDGMENTS
-
CONFLICTS OF INTEREST: No potential conflict of interest relevant to this article was reported.
-
AUTHOR CONTRIBUTIONS:
NOTES
SUPPLEMENTARY MATERIALS
Supplementary Table 1
Supplementary Fig. 1
Supplementary Fig. 2
Supplementary Fig. 3
Supplementary Fig. 4
Supplementary Fig. 5
- 1. Reidy K, Kang HM, Hostetter T, Susztak K. Molecular mechanisms of diabetic kidney disease. J Clin Invest 2014;124:2333-2340. ArticlePubMedPMC
- 2. Gallagher H, Suckling RJ. Diabetic nephropathy: where are we on the journey from pathophysiology to treatment? Diabetes Obes Metab 2016;18:641-647. ArticlePubMedPDF
- 3. Jha JC, Banal C, Okabe J, Gray SP, Hettige T, Chow BSM, Thallas-Bonke V, De Vos L, Holterman CE, Coughlan MT, Power DA, Skene A, Ekinci EI, Cooper ME, Touyz RM, Kennedy CR, Jandeleit-Dahm K. NADPH oxidase Nox5 accelerates renal injury in diabetic nephropathy. Diabetes 2017;66:2691-2703. ArticlePubMedPDF
- 4. Pontrelli P, Conserva F, Papale M, Oranger A, Barozzino M, Vocino G, Rocchetti MT, Gigante M, Castellano G, Rossini M, Simone S, Laviola L, Giorgino F, Grandaliano G, Di Paolo S, Gesualdo L. Lysine 63 ubiquitination is involved in the progression of tubular damage in diabetic nephropathy. FASEB J 2017;31:308-319. ArticlePubMedPDF
- 5. Zhao Y, Yin Z, Li H, Fan J, Yang S, Chen C, Wang DW. MiR-30c protects diabetic nephropathy by suppressing epithelial-to-mesenchymal transition in db/db mice. Aging Cell 2017;16:387-400. ArticlePubMedPMCPDF
- 6. Thiery JP, Acloque H, Huang RY, Nieto MA. Epithelial-mesenchymal transitions in development and disease. Cell 2009;139:871-890. ArticlePubMed
- 7. Higgins SP, Tang Y, Higgins CE, Mian B, Zhang W, Czekay RP, Samarakoon R, Conti DJ, Higgins PJ. TGF-β1/p53 signaling in renal fibrogenesis. Cell Signal 2018;43:1-10. ArticlePubMed
- 8. Ma J, Zhang L, Hao J, Li N, Tang J, Hao L. Up-regulation of microRNA-93 inhibits TGF-β1-induced EMT and renal fibrogenesis by down-regulation of Orai1. J Pharmacol Sci 2018;136:218-227. ArticlePubMed
- 9. Piccolo S. P53 regulation orchestrates the TGF-beta response. Cell 2008;133:767-769. ArticlePubMed
- 10. Yang H, Feng A, Lin S, Yu L, Lin X, Yan X, Lu X, Zhang C. Fibroblast growth factor-21 prevents diabetic cardiomyopathy via AMPK-mediated antioxidation and lipid-lowering effects in the heart. Cell Death Dis 2018;9:227. ArticlePubMedPMCPDF
- 11. Zhang C, Shao M, Yang H, Chen L, Yu L, Cong W, Tian H, Zhang F, Cheng P, Jin L, Tan Y, Li X, Cai L, Lu X. Attenuation of hyperlipidemia- and diabetes-induced early-stage apoptosis and late-stage renal dysfunction via administration of fibroblast growth factor-21 is associated with suppression of renal inflammation. PLoS One 2013;8:e82275. ArticlePubMedPMC
- 12. Li F, Liu Z, Tang C, Cai J, Dong Z. FGF21 is induced in cisplatin nephrotoxicity to protect against kidney tubular cell injury. FASEB J 2018;32:3423-3433. ArticlePubMedPMCPDF
- 13. Wang XM, Xiao H, Liu LL, Cheng D, Li XJ, Si LY. FGF21 represses cerebrovascular aging via improving mitochondrial biogenesis and inhibiting p53 signaling pathway in an AMPK-dependent manner. Exp Cell Res 2016;346:147-156. ArticlePubMed
- 14. Zhang Q, Li Y, Liang T, Lu X, Liu X, Zhang C, Jiang X, Martin RC, Cheng M, Cai L. Loss of FGF21 in diabetic mouse during hepatocellular carcinogenetic transformation. Am J Cancer Res 2015;5:1762-1774. PubMedPMC
- 15. Guo D, Xiao L, Hu H, Liu M, Yang L, Lin X. FGF21 protects human umbilical vein endothelial cells against high glucose-induced apoptosis via PI3K/Akt/Fox3a signaling pathway. J Diabetes Complications 2018;32:729-736. ArticlePubMed
- 16. Zhang C, Zhang L, Chen S, Feng B, Lu X, Bai Y, Liang G, Tan Y, Shao M, Skibba M, Jin L, Li X, Chakrabarti S, Cai L. The prevention of diabetic cardiomyopathy by non-mitogenic acidic fibroblast growth factor is probably mediated by the suppression of oxidative stress and damage. PLoS One 2013;8:e82287. ArticlePubMedPMC
- 17. Shao M, Yu L, Zhang F, Lu X, Li X, Cheng P, Lin X, He L, Jin S, Tan Y, Yang H, Zhang C, Cai L. Additive protection by LDR and FGF21 treatment against diabetic nephropathy in type 2 diabetes model. Am J Physiol Endocrinol Metab 2015;309:E45-E54. ArticlePubMedPMC
- 18. Shao M, Lu X, Cong W, Xing X, Tan Y, Li Y, Li X, Jin L, Wang X, Dong J, Jin S, Zhang C, Cai L. Multiple low-dose radiation prevents type 2 diabetes-induced renal damage through attenuation of dyslipidemia and insulin resistance and subsequent renal inflammation and oxidative stress. PLoS One 2014;9:e92574. ArticlePubMedPMC
- 19. Zhang C, Lu X, Tan Y, Li B, Miao X, Jin L, Shi X, Zhang X, Miao L, Li X, Cai L. Diabetes-induced hepatic pathogenic damage, inflammation, oxidative stress, and insulin resistance was exacerbated in zinc deficient mouse model. PLoS One 2012;7:e49257. ArticlePubMedPMC
- 20. Gu J, Wang B, Liu Y, Zhong L, Tang Y, Guo H, Jiang T, Wang L, Li Y, Cai L. Murine double minute 2 siRNA and wild-type p53 gene therapy interact positively with zinc on prostate tumours in vitro and in vivo. Eur J Cancer 2014;50:1184-1194. ArticlePubMed
- 21. Zhang Z, Wang S, Zhou S, Yan X, Wang Y, Chen J, Mellen N, Kong M, Gu J, Tan Y, Zheng Y, Cai L. Sulforaphane prevents the development of cardiomyopathy in type 2 diabetic mice probably by reversing oxidative stress-induced inhibition of LKB1/AMPK pathway. J Mol Cell Cardiol 2014;77:42-52. ArticlePubMed
- 22. Zhang F, Yu L, Lin X, Cheng P, He L, Li X, Lu X, Tan Y, Yang H, Cai L, Zhang C. Minireview: roles of fibroblast growth factors 19 and 21 in metabolic regulation and chronic diseases. Mol Endocrinol 2015;29:1400-1413. ArticlePubMedPMC
- 23. Anuwatmatee S, Tang S, Wu BJ, Rye KA, Ong KL. Fibroblast growth factor 21 in chronic kidney disease. Clin Chim Acta 2019;489:196-202. ArticlePubMed
- 24. Lee CH, Hui EY, Woo YC, Yeung CY, Chow WS, Yuen MM, Fong CH, Xu A, Lam KS. Circulating fibroblast growth factor 21 levels predict progressive kidney disease in subjects with type 2 diabetes and normoalbuminuria. J Clin Endocrinol Metab 2015;100:1368-1375. ArticlePubMedPDF
- 25. Trakarnvanich T, Prommool S, Kurathong S, Teepprasan T, Wang Y. Associations among cardio-ankle vascular index, carotid intima-media thickness, and fibroblast growth factor-21 levels in kidney transplant patients. Transplant Proc 2017;49:1791-1796. ArticlePubMed
- 26. Bagheri L, Hami M, Mojahedi MJ, Ghorban Sabbagh M, Ayatollahi H. Association of metabolic syndrome with serum fibroblast growth factor 21 in kidney transplanted patients. J Renal Inj Prev 2016;5:79-84. ArticlePubMedPMC
- 27. Kohara M, Masuda T, Shiizaki K, Akimoto T, Watanabe Y, Honma S, Sekiguchi C, Miyazawa Y, Kusano E, Kanda Y, Asano Y, Kuro-O M, Nagata D. Association between circulating fibroblast growth factor 21 and mortality in end-stage renal disease. PLoS One 2017;12:e0178971. ArticlePubMedPMC
- 28. Esteghamati A, Khandan A, Momeni A, Behdadnia A, Ghajar A, Nikdad MS, Noshad S, Nakhjavani M, Afarideh M. Circulating levels of fibroblast growth factor 21 in early-stage diabetic kidney disease. Ir J Med Sci 2017;186:785-794. ArticlePubMedPDF
- 29. Hindricks J, Ebert T, Bachmann A, Kralisch S, Lossner U, Kratzsch J, Stolzenburg JU, Dietel A, Beige J, Anders M, Bast I, Bluher M, Stumvoll M, Fasshauer M. Serum levels of fibroblast growth factor-21 are increased in chronic and acute renal dysfunction. Clin Endocrinol (Oxf) 2014;80:918-924. ArticlePubMed
- 30. Han SH, Choi SH, Cho BJ, Lee Y, Lim S, Park YJ, Moon MK, Lee HK, Kang SW, Han DS, Kim YB, Jang HC, Park KS. Serum fibroblast growth factor-21 concentration is associated with residual renal function and insulin resistance in end-stage renal disease patients receiving long-term peritoneal dialysis. Metabolism 2010;59:1656-1662. ArticlePubMed
- 31. Shi YC, Lu WW, Hou YL, Fu K, Gan F, Cheng SJ, Wang SP, Qi YF, Liu JH. Protection effect of exogenous fibroblast growth factor 21 on the kidney injury in vascular calcification rats. Chin Med J (Engl) 2018;131:532-538. ArticlePubMedPMC
- 32. Kim HW, Lee JE, Cha JJ, Hyun YY, Kim JE, Lee MH, Song HK, Nam DH, Han JY, Han SY, Han KH, Kang YS, Cha DR. Fibroblast growth factor 21 improves insulin resistance and ameliorates renal injury in db/db mice. Endocrinology 2013;154:3366-3376. ArticlePubMedPDF
- 33. Wente W, Efanov AM, Brenner M, Kharitonenkov A, Koster A, Sandusky GE, Sewing S, Treinies I, Zitzer H, Gromada J. Fibroblast growth factor-21 improves pancreatic beta-cell function and survival by activation of extracellular signal-regulated kinase 1/2 and Akt signaling pathways. Diabetes 2006;55:2470-2478. PubMed
- 34. Kalluri R, Weinberg RA. The basics of epithelial-mesenchymal transition. J Clin Invest 2009;119:1420-1428. ArticlePubMedPMC
- 35. Kalluri R. EMT: when epithelial cells decide to become mesenchymal-like cells. J Clin Invest 2009;119:1417-1419. ArticlePubMedPMC
- 36. Tennakoon AH, Izawa T, Kuwamura M, Yamate J. Pathogenesis of type 2 epithelial to mesenchymal transition (EMT) in renal and hepatic fibrosis. J Clin Med 2015;5:E4. Article
- 37. Burns WC, Thomas MC. The molecular mediators of type 2 epithelial to mesenchymal transition (EMT) and their role in renal pathophysiology. Expert Rev Mol Med 2010;12:e17. ArticlePubMed
- 38. Ling L, Tan Z, Zhang C, Gui S, Hu Y, Chen L. Long noncoding RNA ENSRNOG00000037522 is involved in the podocyte epithelial-mesenchymal transition in diabetic rats. Int J Mol Med 2018;41:2704-2714. ArticlePubMedPMC
- 39. Ying Q, Wu G. Molecular mechanisms involved in podocyte EMT and concomitant diabetic kidney diseases: an update. Ren Fail 2017;39:474-483. ArticlePubMedPMCPDF
- 40. Hills CE, Squires PE. The role of TGF-β and epithelial-to mesenchymal transition in diabetic nephropathy. Cytokine Growth Factor Rev 2011;22:131-139. ArticlePubMed
- 41. Wang B, Koh P, Winbanks C, Coughlan MT, McClelland A, Watson A, Jandeleit-Dahm K, Burns WC, Thomas MC, Cooper ME, Kantharidis P. miR-200a Prevents renal fibrogenesis through repression of TGF-β2 expression. Diabetes 2011;60:280-287. ArticlePubMedPDF
- 42. Kalo E, Buganim Y, Shapira KE, Besserglick H, Goldfinger N, Weisz L, Stambolsky P, Henis YI, Rotter V. Mutant p53 attenuates the SMAD-dependent transforming growth factor beta1 (TGF-beta1) signaling pathway by repressing the expression of TGF-beta receptor type II. Mol Cell Biol 2007;27:8228-8242. PubMedPMC
- 43. Kawarada Y, Inoue Y, Kawasaki F, Fukuura K, Sato K, Tanaka T, Itoh Y, Hayashi H. TGF-beta induces p53/Smads complex formation in the PAI-1 promoter to activate transcription. Sci Rep 2016;6:35483. PubMedPMC
- 44. Yan J, Wang J, Huang H, Huang Y, Mi T, Zhang C, Zhang L. Fibroblast growth factor 21 delayed endothelial replicative senescence and protected cells from H(2)O(2)-induced premature senescence through SIRT1. Am J Transl Res 2017;9:4492-4501. PubMedPMC
- 45. Jiang X, Chen J, Zhang C, Zhang Z, Tan Y, Feng W, Skibba M, Xin Y, Cai L. The protective effect of FGF21 on diabetes-induced male germ cell apoptosis is associated with up-regulated testicular AKT and AMPK/Sirt1/PGC-1α signaling. Endocrinology 2015;156:1156-1170. ArticlePubMedPMC
- 46. Purvis GSD, Chiazza F, Chen J, Azevedo-Loiola R, Martin L, Kusters DHM, Reutelingsperger C, Fountoulakis N, Gnudi L, Yaqoob MM, Collino M, Thiemermann C, Solito E. Annexin A1 attenuates microvascular complications through restoration of Akt signalling in a murine model of type 1 diabetes. Diabetologia 2018;61:482-495. ArticlePubMedPDF
- 47. Li B, Cui W, Tan Y, Luo P, Chen Q, Zhang C, Qu W, Miao L, Cai L. Zinc is essential for the transcription function of Nrf2 in human renal tubule cells in vitro and mouse kidney in vivo under the diabetic condition. J Cell Mol Med 2014;18:895-906. ArticlePubMedPMCPDF
- 48. Mayo LD, Donner DB. The PTEN, Mdm2, p53 tumor suppressor-oncoprotein network. Trends Biochem Sci 2002;27:462-467. ArticlePubMed
- 49. Chi SW, Lee SH, Kim DH, Ahn MJ, Kim JS, Woo JY, Torizawa T, Kainosho M, Han KH. Structural details on mdm2-p53 interaction. J Biol Chem 2005;280:38795-38802. ArticlePubMed
- 50. Moll UM, Petrenko O. The MDM2-p53 interaction. Mol Cancer Res 2003;1:1001-1008. PubMed
REFERENCES
Effects of fibroblast growth factor 21 (FGF21) supplement on renal extracellular matrix (ECM) accumulation in diabetic nephropathy (DN) mice. Since ECM leads to renal fibrosis, Western blotting was used to examine the expressions of the markers of ECM and fibrosis in the kidneys, including (A) connective tissue growth factor (CTGF), (B) fibronectin 1 (FN1), (C) collagen I (Col I), (D) collagen II (Col II), (E) collagen IV (Col IV), and (F) plasminogen activator inhibitor-1 (PAI-1). The mRNA level of multiple markers of EMC accumulation and renal fibrosis including (G) CTGF, (H) FN1, (I) Col I, (J) Col II, (K) Col IV, (L) and PAI-1 in the kidney with or without FGF21 treatment were examined by quantitative polymerase chain reaction. Data are presented as the mean±standard deviation (n=8/group). aP<0.05 vs. the control (Con) group, bP<0.05 vs. DN group.
Effects of fibroblast growth factor 21 (FGF21) supplement on the formation and nuclear translocation of p53/Smad2/3 complex. Western blot analysis was used to examine the expression of (A) renal transforming growth factor beta (TGF-β) and phosphorylation of (B) Smad2, (C) Smad3, and (D) Smad7 as well as (E) renal p53 phosphorylation. (F, G) Immunoprecipitation (IP) was used to examine the protein-protein interaction between p53 and Smad2/3 with or without FGF21 treatment. The nuclear translocation of p53/Smad2/3 complex was examined by changes in the expression of nuclear (H) p53, (I) Smad2, (J) and Smad3, as determined by Western blot analysis. Data are presented as the mean±standard deviation (n=8/group). aP<0.05 vs. the control (Con) group, bP<0.05 vs. diabetic nephropathy (DN) group.
Effect of pifithrin-α (PF-α) on fibroblast growth factor 21 (FGF21)-induced suppression of renal p53 phosphorylation and interaction of p53/Smad2/3. (A) The phosphorylation and expression of renal p53 in the presence of FGF21 and/or PF-α were examined by Western blotting. (B, C) The protein-protein interaction between p53 and Smad2/3 in the presence of FGF21 and/or PF-α was examined by immunoprecipitation (IP). Western blot analysis was used to examine the activity and (D) expression of renal AKT and the expression of the extracellular matrix (ECM) and fibrotic markers in the kidneys including (E, F) connective tissue growth factor (CTGF), (E, G) fibronectin 1 (FN1), (E, H) plasminogen activator inhibitor-1 (PAI-1), (E, I) collagen I (Col I), (E, J) collagen II (Col II), and (E, K) collagen IV (Col IV). Data are presented as the mean±standard deviation (n=8/group). aP<0.05 vs. the diabetic nephropathy (DN) group.
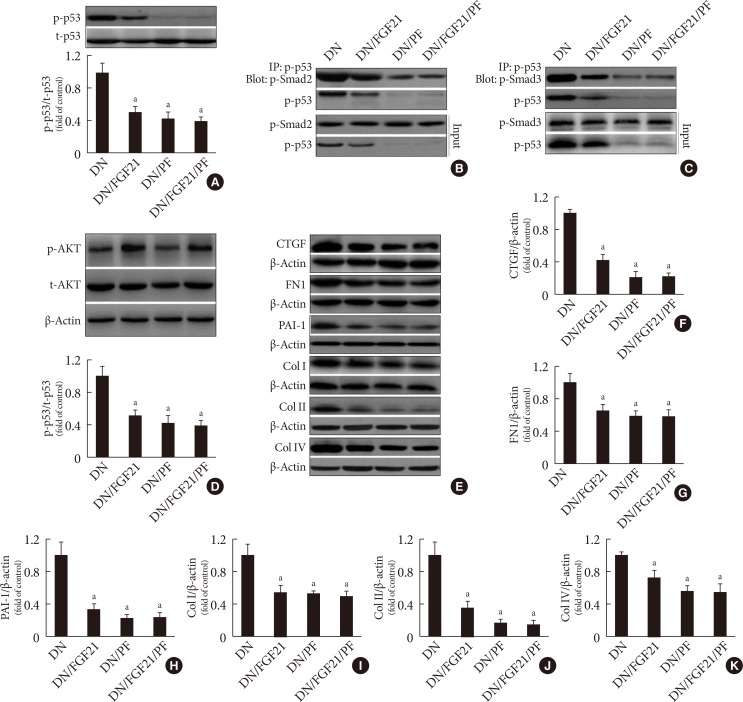
Effects of 10-[4′-(N,N-Diethylamino)butyl]-2-chlorophenoxazine hydrochloride (10-DEBC) on fibroblast growth factor 21 (FGF21)-induced suppression of renal p53 phosphorylation and complex formation of p53/Smad2/3. The phosphorylation and expression of renal (A, B) AKT, (A, C) p53, and (A, D) mouse double minute-2 homolog (MDM2) in the presence of FGF21 and/or 10-DEBC were examined by Western blotting. (E, F) The protein-protein interaction between p53 and Smad2/3 in the presence of FGF21 and/or 10-DEBC was examined by immunoprecipitation (IP). Data are presented as the mean±standard deviation (n=8/group). DEBC=10-DEBC (10-DEBC hydrochloride). aP<0.05 vs. the diabetic nephropathy (DN) group.

Effects of 10-[4′-(N,N-Diethylamino)butyl]-2-chlorophenoxazine hydrochloride (10-DEBC) on fibroblast growth factor 21 (FGF21)-induced prevention of the epithelial-to-mesenchymal transition (EMT) in the diabetic kidney. Western blot analysis was used to examine the markers of EMT and extracellular matrix (ECM) in the kidneys, including (A) E-cadherin, (B) P-cadherin, (C) zonula occludens-1 (ZO-1), (D) α-smooth muscle actin (α-SMA), (E) vimentin, and (F) laminin, (G) connective tissue growth factor (CTGF), (H) fibronectin 1 (FN1), (I) plasminogen activator inhibitor-1 (PAI-1), (J) collagen I (Col I), (K) collagen II (Col II), and (L) collagen IV (Col IV). Data are presented as the mean±standard deviation (n=8/group). aP<0.05 vs. the diabetic nephropathy (DN) group.
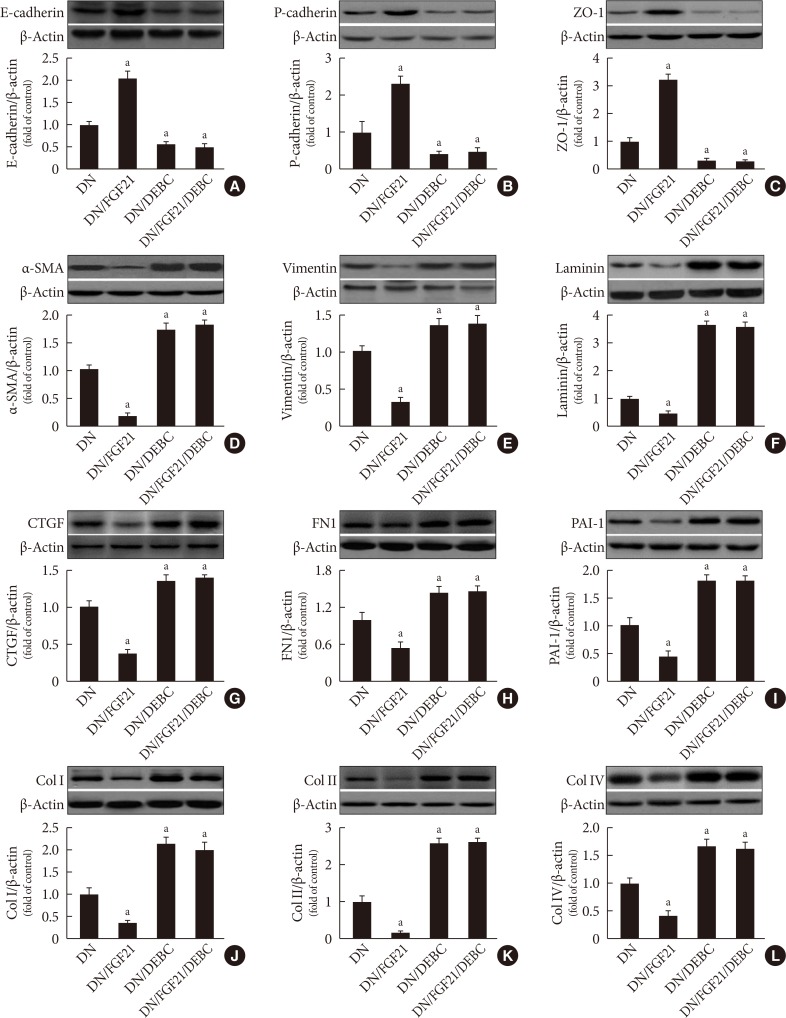
Figure & Data
References
Citations
- Epithelial–mesenchymal plasticity in kidney fibrosis
Sudarat Hadpech, Visith Thongboonkerd
genesis.2024;[Epub] CrossRef - Fibroblast growth factor 21 alleviates unilateral ureteral obstruction-induced renal fibrosis by inhibiting Wnt/β-catenin signaling pathway
Wenhui Zhong, Yuheng Jiang, Huizhen Wang, Xiang Luo, Tao Zeng, Huimi Huang, Ling Xiao, Nan Jia, Aiqing Li
Biochimica et Biophysica Acta (BBA) - Molecular Cell Research.2024; 1871(2): 119620. CrossRef - Platelet concentrates may affect the formation of pathological scars by regulating epithelial to mesenchymal transition
Ju Tian, Dandan Shi, Chenyan Long, Jing Ding, Huimin You, Xiaoying He, Biao Cheng
Medical Hypotheses.2024; 182: 111227. CrossRef - Cadherin-responsive hydrogel combined with dental pulp stem cells and fibroblast growth factor 21 promotes diabetic scald repair via regulating epithelial-mesenchymal transition and necroptosis
Wenjie Lu, Juan Zhao, Xiong Cai, Yutian Wang, Wenwei Lin, Yaoping Fang, Yunyang Wang, Jinglei Ao, Jiahui Shou, Jiake Xu, Sipin Zhu
Materials Today Bio.2024; 24: 100919. CrossRef - Sodium butyrate ameliorated diabetic nephropathy-associated tubulointerstitial inflammation by modulating the tight junctions of renal tubular epithelial cells
Tingting Yang, Lin Li, Cai Heng, Pian Sha, Yiying Wang, Jiaming Shen, Zhenzhou Jiang, Sitong Qian, Chujing Wei, Hao Yang, Xia Zhu, Tao Wang, Mengying Wu, Jianyun Wang, Qian Lu, Xiaoxing Yin
Food & Function.2024; 15(5): 2628. CrossRef - Urinary Excretion of Biomolecules Related to Cell Cycle, Proliferation, and Autophagy in Subjects with Type 2 Diabetes and Chronic Kidney Disease
Anton I. Korbut, Vyacheslav V. Romanov, Vadim V. Klimontov
Biomedicines.2024; 12(3): 487. CrossRef - FGF21 Inhibits Hypoxia/Reoxygenation-induced Renal Tubular Epithelial Cell Injury by Regulating the PPARγ/NF-κB Signaling Pathway
Ruixue Li, Xi Liu
Cell Biochemistry and Biophysics.2024;[Epub] CrossRef - Timosaponin BII inhibits TGF‐β mediated epithelial‐mesenchymal transition through Smad‐dependent pathway during pulmonary fibrosis
Dali Ding, Xuebin Shen, Lizhen Yu, Yueyue Zheng, Yao Liu, Wei Wang, Li Liu, Zitong Zhao, Sihui Nian, Limin Liu
Phytotherapy Research.2023; 37(7): 2787. CrossRef - Fibroblast growth factors (FGFs) endocrines et fibrogenèse pulmonaire
M. Ghanem, A. Mailleux, B. Crestani
Revue des Maladies Respiratoires.2023; 40(3): 239. CrossRef - Mesenchymal Stem Cell Therapy in Kidney Diseases: Potential and Challenges
Fukun Chen, NaNa Chen, Chunjuan Xia, Hongyue Wang, Lishi Shao, Chen Zhou, Jiaping Wang
Cell Transplantation.2023; 32: 096368972311642. CrossRef - MicroRNA functional metal-organic framework nanocomposite for efficient inhibition of drug-resistant breast cancer cells
Yuxin Shen, Yao Zhang, Xiyue Gao, Mengdi Shang, Yanfei Cai, Zhaoqi Yang
Emergent Materials.2023; 6(5): 1537. CrossRef - Downregulation of a potential therapeutic target NPAS2, regulated by p53, alleviates pulmonary fibrosis by inhibiting epithelial-mesenchymal transition via suppressing HES1
Yingying Chen, Zhong He, Bo Zhao, Rui Zheng
Cellular Signalling.2023; 109: 110795. CrossRef - KLF5/MDM2 Axis Modulates Oxidative Stress and Epithelial-Mesenchymal Transition in Human Lens Epithelial Cells: The Role in Diabetic Cataract
Xiao Li, Doudou Chen, Bowen Ouyang, Shengnan Wang, Yawei Li, Li Li, Siquan Zhu, Guangying Zheng
Laboratory Investigation.2023; 103(11): 100226. CrossRef - MAP3K19 Promotes the Progression of Tuberculosis-Induced Pulmonary Fibrosis Through Activation of the TGF-β/Smad2 Signaling Pathway
Yu Xia, Haiyue Wang, Meihua Shao, Xuemei Liu, Feng Sun
Molecular Biotechnology.2023;[Epub] CrossRef - Exploring the pathogenesis of type 2 diabetes mellitus intestinal damp-heat syndrome and the therapeutic effect of Gegen Qinlian Decoction from the perspective of exosomal miRNA
LiSha He, Tingting Bao, Yingying Yang, Han Wang, Chengjuan Gu, Jia Chen, Tiangang Zhai, Xinhui He, Mengyi Wu, Linhua Zhao, Xiaolin Tong
Journal of Ethnopharmacology.2022; 285: 114786. CrossRef - Cardamomin protects from diabetes-induced kidney damage through modulating PI3K/AKT and JAK/STAT signaling pathways in rats
Chan Gao, Xiao Fei, Ming Wang, Qi Chen, Ning Zhao
International Immunopharmacology.2022; 107: 108610. CrossRef - Fibroblast Growth Factor 21 Predicts Short-Term Prognosis in Patients With Acute Heart Failure: A Prospective Cohort Study
Guihai Wu, Shenglin Wu, Jingyi Yan, Shanshan Gao, Jinxiu Zhu, Minghui Yue, Zexin Li, Xuerui Tan
Frontiers in Cardiovascular Medicine.2022;[Epub] CrossRef - Research Progress of Fibroblast Growth Factor 21 in Fibrotic Diseases
Min-Qi Jia, Cha-Xiang Guan, Jia-Hao Tao, Yong Zhou, Liang-Jun Yan
Oxidative Medicine and Cellular Longevity.2022; 2022: 1. CrossRef - Circ_FOXP1 promotes the growth and survival of high glucose-treated human trophoblast cells through the regulation of miR-508-3p/SMAD family member 2 pathway
Mingqun Li, Yuqin Huang, Hongli Xi, Wei Zhang, Ziwu Xiang, Lingyun Wang, Xuanyu Li, Hongyan Guo
Endocrine Journal.2022; 69(9): 1067. CrossRef - Fibroblast growth factor 21 attenuates the progression of hyperuricemic nephropathy through inhibiting inflammation, fibrosis and oxidative stress
Xinghao Jiang, Qing Wu, Yeboah Kwaku Opoku, Yimeng Zou, Dan Wang, Changhui Hu, Guiping Ren
Basic & Clinical Pharmacology & Toxicology.2022; 131(6): 474. CrossRef - Myokines: Novel therapeutic targets for diabetic nephropathy
Ming Yang, Shilu Luo, Jinfei Yang, Wei Chen, Liyu He, Di Liu, Li Zhao, Xi Wang
Frontiers in Endocrinology.2022;[Epub] CrossRef - Dojuksan ameliorates tubulointerstitial fibrosis through irisin-mediated muscle-kidney crosstalk
Songling Jiang, Dal-Seok Oh, Debra Dorotea, Eunjung Son, Dong-Seon Kim, Hunjoo Ha
Phytomedicine.2021; 80: 153393. CrossRef - Chromatin accessibility of kidney tubular cells under stress reveals key transcription factor mediating acute and chronic kidney disease
Yuexian Xing, Qi Wang, Jing Zhang, Wenju Li, Aiping Duan, Jingping Yang, Zhihong Liu
The FEBS Journal.2021; 288(18): 5446. CrossRef - Small molecules against the origin and activation of myofibroblast for renal interstitial fibrosis therapy
Ya-long Feng, Wen-bo Wang, Yue Ning, Hua Chen, Pei Liu
Biomedicine & Pharmacotherapy.2021; 139: 111386. CrossRef - FGF21 prevents low-protein diet-induced renal inflammation in aged mice
Han Fang, Sujoy Ghosh, Landon C. Sims, Kirsten P. Stone, Cristal M. Hill, Denisha Spires, Daria V. Ilatovskaya, Christopher D. Morrison, Thomas W. Gettys, Krisztian Stadler
American Journal of Physiology-Renal Physiology.2021; 321(3): F356. CrossRef - IFN-α-2b Inhibits the Proliferation and Migration of Fibroblasts via the TGFβ/Smad Signaling Pathway to Reduce Postoperative Epidural Fibrosis
Zhendong Liu, Hui Chen, Zhehao Fan, Jihang Dai, Yu Sun, Lianqi Yan, Rui Wang, Xiaolei Li, Jingcheng Wang
Journal of Interferon & Cytokine Research.2021; 41(8): 271. CrossRef - FOXN3 inhibits cell proliferation and invasion via modulating the AKT/MDM2/p53 axis in human glioma
Chaojia Wang, Hanjun Tu, Ling Yang, Chunming Ma, Juntao Hu, Jie Luo, Hui Wang
Aging.2021; 13(17): 21587. CrossRef - Regulation and Potential Biological Role of Fibroblast Growth Factor 21 in Chronic Kidney Disease
Xue Zhou, Yuefeng Zhang, Ning Wang
Frontiers in Physiology.2021;[Epub] CrossRef - The Multiple Roles of Fibroblast Growth Factor in Diabetic Nephropathy
Junyu Deng, Ye Liu, Yiqiu Liu, Wei Li, Xuqiang Nie
Journal of Inflammation Research.2021; Volume 14: 5273. CrossRef - Exercise Training Alleviates Cardiac Fibrosis through Increasing Fibroblast Growth Factor 21 and Regulating TGF-β1-Smad2/3-MMP2/9 Signaling in Mice with Myocardial Infarction
Yixuan Ma, Yixin Kuang, Wenyan Bo, Qiaoqin Liang, Wenfei Zhu, Mengxin Cai, Zhenjun Tian
International Journal of Molecular Sciences.2021; 22(22): 12341. CrossRef - Snai1-induced partial epithelial–mesenchymal transition orchestrates p53–p21-mediated G2/M arrest in the progression of renal fibrosis via NF-κB-mediated inflammation
Ruochen Qi, Jiyan Wang, Yamei Jiang, Yue Qiu, Ming Xu, Ruiming Rong, Tongyu Zhu
Cell Death & Disease.2021;[Epub] CrossRef - Destruction of the blood-retina barrier in diabetic retinopathy depends on angiotensin-converting enzyme-mediated TGF-β1/Smad signaling pathway activation
Ping Sun, Ning Xu, Yan Li, Yang Han
International Immunopharmacology.2020; 85: 106686. CrossRef Chrysophanol Inhibits the Progression of Diabetic Nephropathy via Inactivation of TGF-β Pathway
Chuan Guo, Yarong Wang, Yuanlin Piao, Xiangrong Rao, Dehai Yin
Drug Design, Development and Therapy.2020; Volume 14: 4951. CrossRef
 PubReader
PubReader Cite
Cite