- Current
- Browse
- Collections
-
For contributors
- For Authors
- Instructions to authors
- Article processing charge
- e-submission
- For Reviewers
- Instructions for reviewers
- How to become a reviewer
- Best reviewers
- For Readers
- Readership
- Subscription
- Permission guidelines
- About
- Editorial policy
Articles
- Page Path
- HOME > Diabetes Metab J > Volume 41(2); 2017 > Article
-
ReviewPathophysiology Role of NO/VASP Signaling Pathway against Obesity-Related Inflammation and Insulin Resistance
-
Yu Mi Kang1, Francis Kim2, Woo Je Lee1

-
Diabetes & Metabolism Journal 2017;41(2):89-95.
DOI: https://doi.org/10.4093/dmj.2017.41.2.89
Published online: November 15, 2016
1Department of Internal Medicine, Asan Medical Center, University of Ulsan College of Medicine, Seoul, Korea.
2Department of Medicine, Diabetes and Obesity Center of Excellence, University of Washington, Seattle, WA, USA.
- Corresponding author: Woo Je Lee. Department of Internal Medicine, Asan Medical Center, University of Ulsan College of Medicine, 88 Olympic-ro 43-gil, Songpa-gu, Seoul 05505, Korea. lwjatlas@amc.seoul.kr
• Received: September 8, 2016 • Accepted: October 26, 2016
Copyright © 2017 Korean Diabetes Association
This is an Open Access article distributed under the terms of the Creative Commons Attribution Non-Commercial License (http://creativecommons.org/licenses/by-nc/4.0/) which permits unrestricted non-commercial use, distribution, and reproduction in any medium, provided the original work is properly cited.
ABSTRACT
- Obesity has quickly become a worldwide pandemic, causing major adverse health outcomes such as dyslipidemia, type 2 diabetes mellitus, cardiovascular disease and cancers. Obesity-induced insulin resistance is the key for developing these metabolic disorders, and investigation to understand the molecular mechanisms involved has been vibrant for the past few decades. Of these, low-grade chronic inflammation is suggested as a critical concept in the development of obesity-induced insulin resistance, and the anti-inflammatory effect of nitric oxide (NO) signaling has been reported to be linked to improvement of insulin resistance in multiple organs involved in glucose metabolism. Recently, a body of evidence suggested that vasodilatory-stimulated phosphoprotein (VASP), a downstream mediator of NO signaling plays a crucial role in the anti-inflammatory effect and improvement of peripheral insulin resistance. These preclinical studies suggest that NO/VASP signaling could be an ideal therapeutic target in the treatment of obesity-related metabolic dysfunction. In this review, we introduce studies that investigated the protective role of NO/VASP signaling against obesity-related inflammation and insulin resistance in various tissues.
- Obesity has become a huge burden worldwide. According to the 2013 Korea National Health and Nutrition Examination Survey, one in three Korean adults are obese and more than half are considered to be at least overweight [1], implying that Korea is not an exception to this worldwide pandemic. Environmental factors such as unhealthy dietary habits, sedentary lifestyle and socioeconomic influences are considered as the major contributors.
- In the past several decades, evidence suggests that obesity is a state of chronic, low-grade inflammation. One of the earliest evidence demonstrated the expression of proinflammatory cytokine, tumor necrosis factor α (TNF-α), in rodent adipose tissue during the development of obesity and the attenuation of insulin resistance followed by neutralization of TNF-α [2]. This study supports the role of inflammation in obesity and regulation of its complications by inflammatory mediators [3]. Thereafter, significant advances in understanding the complex role of immune-metabolism have been accomplished, and obesity is now known to be associated with proinflammatory cytokine secretion, immune cell infiltration, and disrupted function of tissues involved in glucose homeostasis, leading to insulin resistance [4].
- Nitric oxide (NO) is an endogenous signaling molecule produced by nitric oxide synthase (NOS). NO is involved in various physiologic processes such as the regulation of synaptic transmission, vasodilation, leukocyte-endothelial interactions, immune function, and angiogenesis [5]. Besides these pleiotropic effects, NO has recently emerged as an important regulator of energy metabolism. For example, NO bioavailability reflected by NOS activity is decreased in animal models of diet-induced obesity (DIO) [6] and in obese and insulin-resistant patients [78]. The reduction of NO content occurs early in the course of developing insulin resistance, followed by vascular and peripheral tissue (such as liver, muscle, and adipose) inflammation and insulin resistance during dietary excess [910]. Furthermore, genetic deletion of endothelial nitric oxide synthase (eNOS) is associated with insulin resistance even in the absence of high-fat (HF) stimuli [11]. Conversely, interventions to increase NO output has remarkable effects on obesity and insulin resistance in both animal and human studies [121314], implying that anti-inflammatory and anti-obesity properties of NO might serve as a potential therapeutic approach against obesity and its related metabolic outcomes.
- Vasodilatory-stimulated phosphoprotein (VASP) is a 40 kDa protein associated with cell adhesion and modulation of cytoskeletons [1415] and found in various cell types, but is highly expressed in vascular endothelial cells, smooth muscle cells, fibroblasts, and platelets [15]. VASP is also known to be a downstream molecule of NO signal transduction pathway. Recently, a growing body of evidence suggests that VASP as a downstream mediator of NO signal transduction pathway exerts anti-obesity effects by improving inflammation and insulin resistance in various tissues involved in glucose metabolism [1011161718]. Therefore, in this review, we evaluate the current evidence supporting the protective effect of NO/cyclic guanosine monophosphate (cGMP)/VASP signaling pathway against obesity and possibly, type 2 diabetes mellitus.
INTRODUCTION
- The eNOS is mainly expressed in endothelial cells and generates eNOS-derived NO [19]. In cardiovascular tissues, eNOS-derived NO is known to exert vasodilatory, anti-inflammatory, and anti-proliferative effects [1920] via cGMP-dependent protein kinase (PKG) by increasing cGMP levels [9]. The NO/cGMP-induced PKG activation in various cell types exerts diverse biological function by modulating its multiple downstream effectors. For example, PKG interacts with VASP [21222324] and survival molecules such as the apoptosis-regulating protein BAD [25], the oncogenic tyrosine kinase c-Src [22], and the transcription factor cyclic adenosine monophosphate (cAMP)-responsive element binding protein (CREB) [26] to regulate cell migration, survival, and proliferation. On the other hand, PKG-mediated activation of the mitogen-activated protein kinase (MAPK) family regulates angiogenesis in vascular endothelial cells [27] and the contractile function of myocardial fibers [28] and vascular smooth muscle cells (Fig. 1) [29].
- Of numerous various downstream effectors of PKG, implication of modulating VASP in cancerous or neural cells has been widely studied due to its role in cell adhesion, migration, and proliferation [21222324]. However, recently, phosphorylation of VASP by PKG on a specific serine residue (Ser-239) has been found to improve inflammation and insulin resistance in various peripheral organs such as the adipose tissue, the liver, and the vasculature [1011161718].
NO/cGMP/VASP SIGNALING PATHWAY
- In visceral adipose tissue, nutrient excess leads to the infiltration of macrophages via the production of monocyte chemoattractant protein 1 (MCP-1), a key mediator recruiting macrophage precursors into the adipose tissue [3031]. Subsequent release of proinflammatory cytokines such as interleukin 6 (IL-6) and TNF-α results in impaired insulin signaling [3]. Hence, improving obesity-induced insulin resistance by regulating inflammatory signaling has been of great interest as it can serve as a potential therapeutic target.
- Based on the anti-inflammatory role of endothelial NO signaling in the vasculature and the decline of vascular NO levels early in the course of DIO, Handa et al. [10] made an attempt to unravel the role of NO/cGMP/VASP signaling pathway in the pathogenesis of obesity-associated inflammation in adipose tissue. In this study, HF feeding significantly reduced phospho-eNOS and phospho-VASP in white adipose tissue, compared to mice fed a low-fat (LF) diet. Genetic deficiency of eNOS induced increased expression of proinflammatory cytokines in adipose tissue of eNOS–/– mice; however, a LF diet could not prevent adipose tissue inflammation in these mice, further reflecting the significance of endothelial NO signaling in attenuating adipose tissue inflammation. By contrast, administration of sildenafil, a drug that increases signaling downstream of vascular NO, prevented HF diet-induced proinflammatory gene expression and adipose tissue macrophage infiltration while improving insulin sensitivity in wild type (WT) mice [10].
- Because the authors hypothesized that VASP, a downstream target of the NO/cGMP pathway, is crucial to the anti-inflammatory effects of NO signaling pathway on adipose inflammation, they used VASP–/– and WT littermate mice fed an LF and HF diet [10]. As expected, in VASP–/– mice, levels of mRNA encoding proinflammatory cytokines MCP-1, IL-6, and TNF-α were markedly elevated compared with WT controls [10]. The recapitulation of the effect of eNOS deficiency to increase adipose tissue inflammation in VASP-null mice suggests that VASP must be present for NO/cGMP signaling pathway to exert its anti-inflammatory effect in adipose tissue.
NO/VASP SIGNALING IN ADIPOSE TISSUE
- The liver is a pivotal player in glucose metabolism and inflammatory responses [32]. It is highly vascularized receiving 20% of cardiac output, and hepatic sinusoidal endothelial cells make up half of nonparenchymal cells of the liver [33]. Sinusoidal endothelial cells have shown to express eNOS and produce NO basally, regulating hepatic vascular resistance [34]. In a DIO setting, reduced vascular NO levels were followed by enhanced liver nuclear factor-κB (NF-κB) signaling and impaired insulin-mediated phosphorylation of Akt [35], highlighting the possible role of NO signaling in regulation of hepatic insulin resistance and inflammation.
- Macrophages are myeloid-derived mononuclear cells that are a critical component of the innate immune response. They are more enriched in the areas that are exposed to pathogens, toxins and tissue damages such as the lung and the liver. Accumulating evidence supports a role for tissue macrophages in a broad spectrum of inflammatory conditions [36], and the number and activity of macrophages are associated with insulin resistance and metabolic deterioration in states of over nutrition such as obesity [3738].
- Kupffer cells, the liver-specific macrophages, reside on the luminal side of the sinusoidal endothelium, and are mainly involved in scavenging, host immunity and immune tolerance in the liver [39]. Recent studies demonstrated that increased Kupffer cell activity is responsible for hepatic insulin resistance [4041], and that depletion of Kupffer cells attenuated the development of hepatic steatosis and hepatic insulin resistance, both of which suggest an important early role of Kupffer cells in diet-induced alterations in hepatic insulin resistance [19].
- Tateya et al. [11] questioned whether endothelial NO signaling contributes to the effect of HF feeding to induce inflammatory activation of Kupffer cells and associated hepatic insulin resistance. In the study, proinflammatory activation of Kupffer cells was induced both in WT mice fed HF diet and in genetically eNOS-deficient mice, highlighting the protective effect of NO signaling in hepatic inflammation and insulin resistance [11]. Similarly, targeted deletion of VASP in vivo, a key downstream target of endothelially derived NO, predisposed to hepatic and Kupffer cell inflammation regardless of diet, whereas agent-enhanced VASP signaling reversed the inflammatory activity and insulin resistance in VASP-deficient hepatocytes and macrophages in vitro [11]. Subsequently, Tateya et al. [18] have also demonstrated that NO/VASP signaling increases hepatic fatty acid oxidation by activating AMPK in mice models, suggesting that direct activation of VASP could be a potential therapeutic target of hepatic steatosis.
- The mechanism through which endothelial NO/cGMP/VASP signaling exerts protective effect against inflammation and insulin resistance in Kupffer cells has also been explored [17]. Using mice model with transgenic eNOS overexpression, this study demonstrated that the protective effect of NO signaling against HF diet-induced hepatic inflammation and insulin resistance is associated with reduced proinflammatory M1 and increased anti-inflammatory M2 activation of Kupffer cells [17]. Similar effects were induced by overexpression of VASP in macrophages, whereas VASP deficiency induced proinflammatory M1 macrophage activation. In attempt to determine whether VASP deficiency, specifically in the macrophage compartment, is sufficient to explain the hepatic inflammation and hepatic insulin resistance, transplantation of bone marrow from VASP-deficient donor mice into normal recipients was performed. As a result, the transplantation led to hepatic inflammation and insulin resistance resembling that induced in normal mice fed HF diet [17]. Taken together, NO/VASP signal transduction inhibits proinflammatory M1 activation of the Kupffer cells, and it can serve as a physiological determinant of macrophage polarization and a promising therapeutic target to prevent hepatic inflammation and insulin resistance.
NO/VASP SIGNALING IN THE LIVER
- Metabolic deterioration related to obesity is linked with cardiovascular diseases, and atherosclerosis is responsible for the vast majority of these cardiovascular events. Thus, it is important to detect and delay the progression of atherosclerosis in its early stage. In recent years, it has become evident that insulin resistance and endothelial dysfunction play a central role in the pathogenesis of atherosclerosis [42]. Metabolic dysfunction causes lipid deposition and oxidative stress to the vessel wall, triggering an inflammatory reaction, and the release of chemoattractants and cytokines worsens the insulin resistance and endothelial dysfunction [4243]. Within this context, therapies that improve vascular insulin resistance and inflammation would be ideal as it may reduce cardiovascular morbidity and mortality in clinical settings.
- Reduced NO bioavailability occurs within 1 week of HF feeding in mice, causing endothelial cell to be significantly more vulnerable to the inflammatory effects of excess nutrition [35]. This further reduces NO production, leading to a "vicious cycle" of increased vascular inflammation and reduced NO levels [16]. By contrast, increasing downstream NO signaling in mice fed HF diet attenuates vascular inflammation [44]; thereby, breaking this vicious cycle and restoring vascular insulin sensitivity [16].
- Cheng et al. [16] investigated the role of VASP as a downstream mediator of the anti-inflammatory effect of NO signaling in vascular tissue. Compared to mice fed a LF diet, markedly reduced levels of VASP Ser239 phosphorylation, a marker of VASP activation, were observed in aortic tissue of DIO mice. HF feeding was associated with increased aortic inflammation, as measured by increased NF-κB dependent gene expression, and reduced vascular insulin sensitivity (including insulin-stimulated phosphorylation of eNOS and Akt) [16]. These HF-diet-induced responses recapitulated in VASP–/– mice, whereas overexpression of VASP in endothelial cells blocked inflammation and insulin resistance induced by palmitate, reflecting the protective effect against inflammatory signaling and insulin resistance in vasculature [16]. These findings implicate that VASP can serve as a potential therapeutic target in the treatment of obesity-related vascular inflammation and insulin resistance.
NO/VASP SIGNALING IN VASCULAR ENDOTHELIUM
- Peripheral insulin resistance in obese state is crucial to the etiology of metabolic disorders such as type 2 diabetes mellitus [45], and thus, understanding the molecular mechanisms of insulin resistance in the organs involved in glucose homeostasis can provide further insights to the development of potential targeted therapy. The series of studies we have evaluated in this review commonly indicate that NO/cGMP signaling limits obesity-related inflammation and insulin resistance in multiple organs, such as adipose tissue, liver, and vascular endothelium and also that VASP is a critical downstream mediator required for this protective effect of NO/cGMP signaling (Fig. 2). Developing potential drugs that alter NO signaling has been of caution due to its cytotoxic effect at high levels, generated by inducible NOS. However, targeting downstream molecules of NO-cGMP signaling pathway, such as soluble guanylyl cyclase or PKG could potentially overcome this drawback NO-targeted therapeutic approach possesses [10]. In this standpoint, VASP can be a promising therapeutic target to limit peripheral insulin resistance in obese state, although accumulation of more evidence is necessitated.
CONCLUSIONS
-
CONFLICTS OF INTEREST: No potential conflict of interest relevant to this article was reported.
NOTES
- 1. Korean Diabetes Association. Korean Diabetes Fact Sheet 2015. Seoul: Korean Diabetes Association; 2015. p. 1-28.
- 2. Hotamisligil GS, Shargill NS, Spiegelman BM. Adipose expression of tumor necrosis factor-alpha: direct role in obesity-linked insulin resistance. Science 1993;259:87-91. ArticlePubMed
- 3. Rocha VZ, Libby P. Obesity, inflammation, and atherosclerosis. Nat Rev Cardiol 2009;6:399-409. ArticlePubMedPDF
- 4. McArdle MA, Finucane OM, Connaughton RM, McMorrow AM, Roche HM. Mechanisms of obesity-induced inflammation and insulin resistance: insights into the emerging role of nutritional strategies. Front Endocrinol (Lausanne) 2013;4:52ArticlePubMedPMC
- 5. Sansbury BE, Hill BG. Regulation of obesity and insulin resistance by nitric oxide. Free Radic Biol Med 2014;73:383-399. ArticlePubMed
- 6. Bender SB, Herrick EK, Lott ND, Klabunde RE. Diet-induced obesity and diabetes reduce coronary responses to nitric oxide due to reduced bioavailability in isolated mouse hearts. Diabetes Obes Metab 2007;9:688-696. ArticlePubMed
- 7. Gruber HJ, Mayer C, Mangge H, Fauler G, Grandits N, Wilders-Truschnig M. Obesity reduces the bioavailability of nitric oxide in juveniles. Int J Obes (Lond) 2008;32:826-831. ArticlePubMedPDF
- 8. Higashi Y, Sasaki S, Nakagawa K, Matsuura H, Chayama K, Oshima T. Effect of obesity on endothelium-dependent, nitric oxide-mediated vasodilation in normotensive individuals and patients with essential hypertension. Am J Hypertens 2001;14:1038-1045. ArticlePubMed
- 9. Francis SH, Busch JL, Corbin JD, Sibley D. cGMP-dependent protein kinases and cGMP phosphodiesterases in nitric oxide and cGMP action. Pharmacol Rev 2010;62:525-563. ArticlePubMedPMC
- 10. Handa P, Tateya S, Rizzo NO, Cheng AM, Morgan-Stevenson V, Han CY, Clowes AW, Daum G, O'Brien KD, Schwartz MW, Chait A, Kim F. Reduced vascular nitric oxide-cGMP signaling contributes to adipose tissue inflammation during high-fat feeding. Arterioscler Thromb Vasc Biol 2011;31:2827-2835. ArticlePubMedPMC
- 11. Tateya S, Rizzo NO, Handa P, Cheng AM, Morgan-Stevenson V, Daum G, Clowes AW, Morton GJ, Schwartz MW, Kim F. Endothelial NO/cGMP/VASP signaling attenuates Kupffer cell activation and hepatic insulin resistance induced by high-fat feeding. Diabetes 2011;60:2792-2801. ArticlePubMedPMCPDF
- 12. Larsen FJ, Schiffer TA, Borniquel S, Sahlin K, Ekblom B, Lundberg JO, Weitzberg E. Dietary inorganic nitrate improves mitochondrial efficiency in humans. Cell Metab 2011;13:149-159. ArticlePubMed
- 13. Lundberg JO, Carlstrom M, Larsen FJ, Weitzberg E. Roles of dietary inorganic nitrate in cardiovascular health and disease. Cardiovasc Res 2011;89:525-532. ArticlePubMed
- 14. Roberts CK, Vaziri ND, Barnard RJ. Effect of diet and exercise intervention on blood pressure, insulin, oxidative stress, and nitric oxide availability. Circulation 2002;106:2530-2532. ArticlePubMed
- 15. Price CJ, Brindle NP. Vasodilator-stimulated phosphoprotein is involved in stress-fiber and membrane ruffle formation in endothelial cells. Arterioscler Thromb Vasc Biol 2000;20:2051-2056. ArticlePubMed
- 16. Cheng AM, Rizzo-DeLeon N, Wilson CL, Lee WJ, Tateya S, Clowes AW, Schwartz MW, Kim F. Vasodilator-stimulated phosphoprotein protects against vascular inflammation and insulin resistance. Am J Physiol Endocrinol Metab 2014;307:E571-E579. ArticlePubMedPMC
- 17. Lee WJ, Tateya S, Cheng AM, Rizzo-DeLeon N, Wang NF, Handa P, Wilson CL, Clowes AW, Sweet IR, Bomsztyk K, Schwartz MW, Kim F. M2 macrophage polarization mediates anti-inflammatory effects of endothelial nitric ixide signaling. Diabetes 2015;64:2836-2846. PubMedPMC
- 18. Tateya S, Rizzo-De Leon N, Handa P, Cheng AM, Morgan-Stevenson V, Ogimoto K, Kanter JE, Bornfeldt KE, Daum G, Clowes AW, Chait A, Kim F. VASP increases hepatic fatty acid oxidation by activating AMPK in mice. Diabetes 2013;62:1913-1922. ArticlePubMedPMCPDF
- 19. Huang PL. eNOS, metabolic syndrome and cardiovascular disease. Trends Endocrinol Metab 2009;20:295-302. ArticlePubMedPMC
- 20. Liu VW, Huang PL. Cardiovascular roles of nitric oxide: a review of insights from nitric oxide synthase gene disrupted mice. Cardiovasc Res 2008;77:19-29. PubMedPMC
- 21. Kwiatkowski AV, Gertler FB, Loureiro JJ. Function and regulation of Ena/VASP proteins. Trends Cell Biol 2003;13:386-392. ArticlePubMed
- 22. Leung EL, Wong JC, Johlfs MG, Tsang BK, Fiscus RR. Protein kinase G type Ialpha activity in human ovarian cancer cells significantly contributes to enhanced Src activation and DNA synthesis/cell proliferation. Mol Cancer Res 2010;8:578-591. PubMed
- 23. Wong JC, Fiscus RR. Protein kinase G activity prevents pathological-level nitric oxide-induced apoptosis and promotes DNA synthesis/cell proliferation in vascular smooth muscle cells. Cardiovasc Pathol 2010;19:e221-e231. ArticlePubMed
- 24. Wong JC, Fiscus RR. Essential roles of the nitric oxide (no)/cGMP/protein kinase G type-Ialpha (PKG-Ialpha) signaling pathway and the atrial natriuretic peptide (ANP)/cGMP/PKG-Ialpha autocrine loop in promoting proliferation and cell survival of OP9 bone marrow stromal cells. J Cell Biochem 2011;112:829-839. ArticlePubMed
- 25. Johlfs MG, Fiscus RR. Protein kinase G type-Ialpha phosphorylates the apoptosis-regulating protein Bad at serine 155 and protects against apoptosis in N1E-115 cells. Neurochem Int 2010;56:546-553. PubMed
- 26. Wong JC, Bathina M, Fiscus RR. Cyclic GMP/protein kinase G type-Ialpha (PKG-Ialpha) signaling pathway promotes CREB phosphorylation and maintains higher c-IAP1, livin, survivin, and Mcl-1 expression and the inhibition of PKG-Ialpha kinase activity synergizes with cisplatin in non-small cell lung cancer cells. J Cell Biochem 2012;113:3587-3598. PubMedPDF
- 27. Kawasaki K, Smith RS Jr, Hsieh CM, Sun J, Chao J, Liao JK. Activation of the phosphatidylinositol 3-kinase/protein kinase Akt pathway mediates nitric oxide-induced endothelial cell migration and angiogenesis. Mol Cell Biol 2003;23:5726-5737. ArticlePubMedPMCPDF
- 28. Kirk JA, Holewinski RJ, Crowgey EL, Van Eyk JE. Protein kinase G signaling in cardiac pathophysiology: impact of proteomics on clinical trials. Proteomics 2016;16:894-905. ArticlePubMedPDF
- 29. Yan C, Kim D, Aizawa T, Berk BC. Functional interplay between angiotensin II and nitric oxide: cyclic GMP as a key mediator. Arterioscler Thromb Vasc Biol 2003;23:26-36. PubMed
- 30. Kanda H, Tateya S, Tamori Y, Kotani K, Hiasa K, Kitazawa R, Kitazawa S, Miyachi H, Maeda S, Egashira K, Kasuga M. MCP-1 contributes to macrophage infiltration into adipose tissue, insulin resistance, and hepatic steatosis in obesity. J Clin Invest 2006;116:1494-1505. ArticlePubMedPMC
- 31. Westerbacka J, Corner A, Kolak M, Makkonen J, Turpeinen U, Hamsten A, Fisher RM, Yki-Jarvinen H. Insulin regulation of MCP-1 in human adipose tissue of obese and lean women. Am J Physiol Endocrinol Metab 2008;294:E841-E845. ArticlePubMed
- 32. Arkan MC, Hevener AL, Greten FR, Maeda S, Li ZW, Long JM, Wynshaw-Boris A, Poli G, Olefsky J, Karin M. IKK-beta links inflammation to obesity-induced insulin resistance. Nat Med 2005;11:191-198. ArticlePubMedPDF
- 33. Ishibashi H, Nakamura M, Komori A, Migita K, Shimoda S. Liver architecture, cell function, and disease. Semin Immunopathol 2009;31:399-409. ArticlePubMedPDF
- 34. Shah V, Haddad FG, Garcia-Cardena G, Frangos JA, Mennone A, Groszmann RJ, Sessa WC. Liver sinusoidal endothelial cells are responsible for nitric oxide modulation of resistance in the hepatic sinusoids. J Clin Invest 1997;100:2923-2930. ArticlePubMedPMC
- 35. Kim F, Pham M, Maloney E, Rizzo NO, Morton GJ, Wisse BE, Kirk EA, Chait A, Schwartz MW. Vascular inflammation, insulin resistance, and reduced nitric oxide production precede the onset of peripheral insulin resistance. Arterioscler Thromb Vasc Biol 2008;28:1982-1988. ArticlePubMedPMC
- 36. Chawla A, Nguyen KD, Goh YP. Macrophage-mediated inflammation in metabolic disease. Nat Rev Immunol 2011;11:738-749. ArticlePubMedPMCPDF
- 37. Cancello R, Henegar C, Viguerie N, Taleb S, Poitou C, Rouault C, Coupaye M, Pelloux V, Hugol D, Bouillot JL, Bouloumie A, Barbatelli G, Cinti S, Svensson PA, Barsh GS, Zucker JD, Basdevant A, Langin D, Clement K. Reduction of macrophage infiltration and chemoattractant gene expression changes in white adipose tissue of morbidly obese subjects after surgery-induced weight loss. Diabetes 2005;54:2277-2286. ArticlePubMedPDF
- 38. Xu H, Barnes GT, Yang Q, Tan G, Yang D, Chou CJ, Sole J, Nichols A, Ross JS, Tartaglia LA, Chen H. Chronic inflammation in fat plays a crucial role in the development of obesity-related insulin resistance. J Clin Invest 2003;112:1821-1830. ArticlePubMedPMC
- 39. Braet F, Wisse E. Structural and functional aspects of liver sinusoidal endothelial cell fenestrae: a review. Comp Hepatol 2002;1:1ArticlePubMedPMC
- 40. Lanthier N, Molendi-Coste O, Horsmans Y, van Rooijen N, Cani PD, Leclercq IA. Kupffer cell activation is a causal factor for hepatic insulin resistance. Am J Physiol Gastrointest Liver Physiol 2010;298:G107-G116. ArticlePubMed
- 41. Papackova Z, Palenickova E, Dankova H, Zdychova J, Skop V, Kazdova L, Cahova M. Kupffer cells ameliorate hepatic insulin resistance induced by high-fat diet rich in monounsaturated fatty acids: the evidence for the involvement of alternatively activated macrophages. Nutr Metab (Lond) 2012;9:22ArticlePubMedPMCPDF
- 42. Cersosimo E, DeFronzo RA. Insulin resistance and endothelial dysfunction: the road map to cardiovascular diseases. Diabetes Metab Res Rev 2006;22:423-436. ArticlePubMed
- 43. Wisse BE, Kim F, Schwartz MW. Physiology. An integrative view of obesity. Science 2007;318:928-929. ArticlePubMed
- 44. Rizzo NO, Maloney E, Pham M, Luttrell I, Wessells H, Tateya S, Daum G, Handa P, Schwartz MW, Kim F. Reduced NO-cGMP signaling contributes to vascular inflammation and insulin resistance induced by high-fat feeding. Arterioscler Thromb Vasc Biol 2010;30:758-765. ArticlePubMedPMC
- 45. Lyssenko V, Almgren P, Anevski D, Perfekt R, Lahti K, Nissen M, Isomaa B, Forsen B, Homstrom N, Saloranta C, Taskinen MR, Groop L, Tuomi T. Botnia study group. Predictors of and longitudinal changes in insulin sensitivity and secretion preceding onset of type 2 diabetes. Diabetes 2005;54:166-174. ArticlePubMedPDF
REFERENCES
Fig. 1
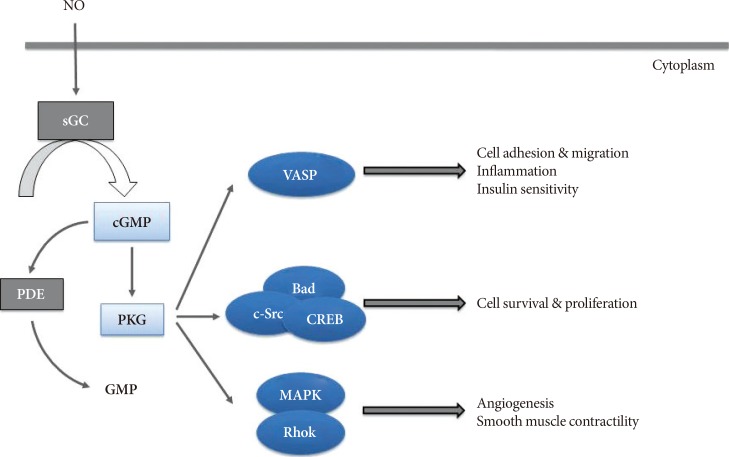
Schematic illustration of nitric oxide (NO)/cyclic guanosine monophosphate (cGMP) signaling pathway and its downstream effects. Activated cGMP-dependent protein kinase (PKG) by NO/cGMP signaling exerts diverse biological function by modulating its multiple downstream effectors in various cell types. PKG interacts with vasodilatory-stimulated phosphoprotein (VASP) to regulate cell adhesion and migration, inflammation and insulin sensitivity, as well as with survival molecules to regulate cell survival, and proliferation. PKG also exerts its role in angiogenesis and smooth muscle contractility via mitogen-activated protein kinase (MAPK) family. sGC, soluble guanylyl cyclase; PDE, phosphodiesterase; c-Src, proto-oncogene tyrosine-protein kinase c-Src; CREB, cyclic adenosine monophosphate (cAMP)-responsive element binding protein; RhoK, rho-associated protein kinase.
Fig. 2
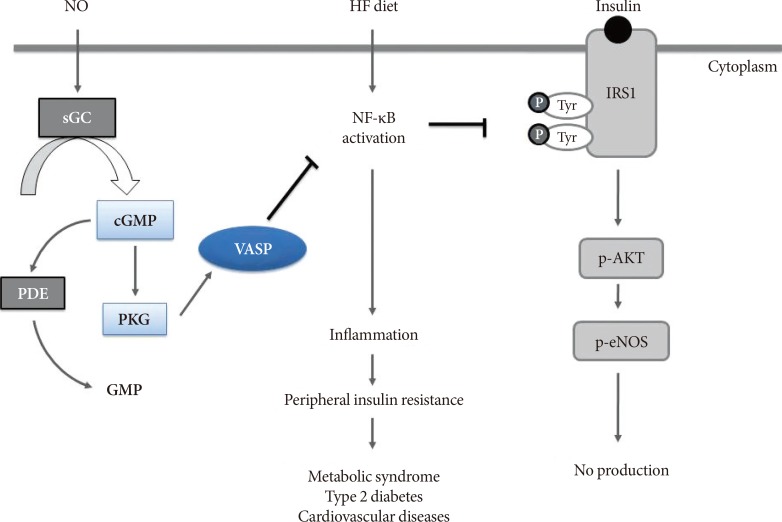
Proposed effect of vasodilatory-stimulated phosphoprotein (VASP) on insulin resistance and inflammatory signaling in multiple organs. Nitric oxide (NO)/cyclic guanosine monophosphate (cGMP) pathway activates VASP, which inhibits nuclear factor-κB (NF-κB) activation; thereby, further inhibiting downstream inflammatory processes and enhancing insulin sensitivity. The schematic end-result of NO/cGMP/VASP signaling suggests its role against cardiometabolic disorders such as metabolic syndrome, type 2 diabetes mellitus, and cardiovascular diseases. Adapted from Cheng et al. [16], with permission from The American Physiological Society. HF, high-fat; sGC, soluble guanylyl cyclase; PDE, phosphodiesterase; PKG, cGMP-dependent protein kinase; IRS1, insulin receptor substrate 1; p-AKT, phospho-Akt; p-eNOS, phospho-endothelial nitric oxide synthase.
Figure & Data
References
Citations
Citations to this article as recorded by 
- Hippocampal proteomic changes in high-fat diet-induced obese mice associated with memory decline
Ping Lu, Cun-Xiu Gao, Fei-Jian Luo, Yu-Ting Huang, Mei-Mei Gao, Yue-Sheng Long
The Journal of Nutritional Biochemistry.2024; 125: 109554. CrossRef - Leptina, obesidad y enfermedades cardiovasculares
Jorly Mejia-Montilla , Nadia Reyna-Villasmil, Andreina Fernández-Ramírez, Eduardo Reyna-Villasmil
Revista Repertorio de Medicina y Cirugía.2023; 32(3): 218. CrossRef - Urinary and circulatory netrin-1 as biomarker in diabetes and its related complications: a systematic review and meta-analysis
Amir Hossein Behnoush, Amirmohammad Khalaji, Zahra Shokri Varniab, Afshin Rahbarghazi, Elahe Amini, Aleksandra Klisic
Endocrine.2023;[Epub] CrossRef - Impact of ultraviolet radiation on cardiovascular and metabolic disorders: The role of nitric oxide and vitamin D
Qing‐Ling Quan, Kyeong‐No Yoon, Ji Su Lee, Eun Ju Kim, Dong Hun Lee
Photodermatology, Photoimmunology & Photomedicine.2023; 39(6): 573. CrossRef - Beneficial Metabolic Effects of Praliciguat, a Soluble Guanylate Cyclase Stimulator, in a Mouse Diet-Induced Obesity Model
Chad D. Schwartzkopf, John R. Hadcock, Guang Liu, Peter Germano, Julien Roux, Courtney M. Shea, Emmanuel S. Buys, Juli E. Jones
Frontiers in Pharmacology.2022;[Epub] CrossRef - Type 2 Diabetes Complicated With Heart Failure: Research on Therapeutic Mechanism and Potential Drug Development Based on Insulin Signaling Pathway
Hui Ye, Yanan He, Chuan Zheng, Fang Wang, Ming Yang, Junzhi Lin, Runchun Xu, Dingkun Zhang
Frontiers in Pharmacology.2022;[Epub] CrossRef - Effect of low-dose tadalafil once daily on glycemic control in patients with type 2 diabetes and erectile dysfunction: a randomized, double-blind, placebo-controlled pilot study
Min-Kyung Lee, Jae-Hyuk Lee, Seo-Young Sohn, Seo Yeon Lee, Tae-Yoong Jeong, Sae Chul Kim
Diabetology & Metabolic Syndrome.2022;[Epub] CrossRef - In situ hydrogel capturing nitric oxide microbubbles accelerates the healing of diabetic foot
Yingzheng Zhao, Lanzi Luo, Lantian Huang, Yingying Zhang, Mengqi Tong, Hanxiao Pan, Jianxun Shangguan, Qing Yao, Shihao Xu, Helin Xu
Journal of Controlled Release.2022; 350: 93. CrossRef - Amelioration effect of black seed oil against high‐fat diet‐induced obesity in rats through Nrf2/HO‐1 pathway
Nada F. Abo El‐Magd, Mohamed El‐Mesery, Amro El‐Karef, Mamdouh M. El‐Shishtawy
Journal of Food Biochemistry.2021;[Epub] CrossRef - An exploratory, randomised, placebo-controlled, 14 day trial of the soluble guanylate cyclase stimulator praliciguat in participants with type 2 diabetes and hypertension
John P. Hanrahan, Jelena P. Seferovic, James D. Wakefield, Phebe J. Wilson, Jennifer G. Chickering, Joon Jung, Kenneth E. Carlson, Daniel P. Zimmer, Andrew L. Frelinger, Alan D. Michelson, Linda Morrow, Michael Hall, Mark G. Currie, G. Todd Milne, Albert
Diabetologia.2020; 63(4): 733. CrossRef - Advantages of Phosphodiesterase Type 5 Inhibitors in the Management of Glucose Metabolism Disorders: A Clinical and Translational Issue
Cristina Antinozzi, Paolo Sgrò, Luigi Di Luigi
International Journal of Endocrinology.2020; 2020: 1. CrossRef - Association of endothelial dysfunction with incident prediabetes, type 2 diabetes and related traits: the KORA F4/FF4 study
Marie-Theres Huemer, Cornelia Huth, Florian Schederecker, Stefanie J Klug, Christa Meisinger, Wolfgang Koenig, Wolfgang Rathmann, Annette Peters, Barbara Thorand
BMJ Open Diabetes Research & Care.2020; 8(1): e001321. CrossRef - Embelin from Embelia ribes ameliorates oxidative stress and inflammation in high-fat diet-fed obese C57BL/6 mice
Priyanka Bansal, Uma Bhandari, Sayeed Ahmad
Pharmacognosy Magazine.2020; 16(5): 443. CrossRef - Weight change is significantly associated with risk of thyroid cancer: A nationwide population-based cohort study
Hyemi Kwon, Kyung-Do Han, Cheol-Young Park
Scientific Reports.2019;[Epub] CrossRef - Diabetes and Cancer: Cancer Should Be Screened in Routine Diabetes Assessment
Sunghwan Suh, Kwang-Won Kim
Diabetes & Metabolism Journal.2019; 43(6): 733. CrossRef - Antioxidant, antihyperglycemic, and antidiabetic activity of Apis mellifera bee tea
Janielle da Silva Melo da Cunha, Tamaeh Monteiro Alfredo, Jéssica Maurino dos Santos, Valter Vieira Alves Junior, Luiza Antas Rabelo, Emerson Silva Lima, Ana Paula de Araújo Boleti, Carlos Alexandre Carollo, Edson Lucas dos Santos, Kely de Picoli Souza, M
PLOS ONE.2018; 13(6): e0197071. CrossRef - Relationship Between Circulating Netrin-1 Concentration, Impaired Fasting Glucose, and Newly Diagnosed Type 2 Diabetes
Jisook Yim, Gyuri Kim, Byung-Wan Lee, Eun Seok Kang, Bong-Soo Cha, Jeong-Ho Kim, Jin Won Cho, Sang-Guk Lee, Yong-ho Lee
Frontiers in Endocrinology.2018;[Epub] CrossRef - Glycyrrhizin ameliorates high fat diet-induced obesity in rats by activating NrF2 pathway
Nada F. Abo El-Magd, Mohamed El-Mesery, Amro El-Karef, Mamdouh M. El-Shishtawy
Life Sciences.2018; 193: 159. CrossRef - cGMP-dependent protein kinase I (cGKI) modulates human hepatic stellate cell activation
Andras Franko, Marketa Kovarova, Susanne Feil, Robert Feil, Robert Wagner, Martin Heni, Alfred Königsrainer, Marc Ruoß, Andreas K. Nüssler, Cora Weigert, Hans-Ulrich Häring, Stefan Z. Lutz, Andreas Peter
Metabolism.2018; 88: 22. CrossRef
 PubReader
PubReader Cite
Cite