- Current
- Browse
- Collections
-
For contributors
- For Authors
- Instructions to authors
- Article processing charge
- e-submission
- For Reviewers
- Instructions for reviewers
- How to become a reviewer
- Best reviewers
- For Readers
- Readership
- Subscription
- Permission guidelines
- About
- Editorial policy
Articles
- Page Path
- HOME > Diabetes Metab J > Volume 47(4); 2023 > Article
-
Original ArticleBasic Research Pharmacologic Activation of Angiotensin-Converting Enzyme II Alleviates Diabetic Cardiomyopathy in db/db Mice by Reducing Reactive Oxidative Stress
-
Donghyun Kim1*
 , Wooju Jeong1*, Yumin Kim1, Jibeom Lee1, Sung Woo Cho2,3, Chang-Myung Oh1, Raekil Park1
, Wooju Jeong1*, Yumin Kim1, Jibeom Lee1, Sung Woo Cho2,3, Chang-Myung Oh1, Raekil Park1 -
Diabetes & Metabolism Journal 2023;47(4):487-499.
DOI: https://doi.org/10.4093/dmj.2022.0125
Published online: April 25, 2023
- 2,199 Views
- 149 Download
1Department of Biomedical Science and Engineering, Gwangju Institute of Science and Technology, Gwangju, Korea
2Division of Cardiology, Department of Internal Medicine, Inje University Ilsan Paik Hospital, College of Medicine, Inje University, Goyang, Korea
3Cardiovascular and Metabolic Disease Center, Smart Marine Therapeutics Center, Inje University, Busan, Korea
- Corresponding authors: Sung Woo Cho https://orcid.org/0000-0003-3983-817X Division of Cardiology, Department of Internal Medicine, Inje University Ilsan Paik Hospital, College of Medicine, Inje University, 170 Juhwa-ro, Ilsanseo-gu, Goyang 10380, Korea E-mail: drswcho@hanmail.net
-
Chang-Myung Oh Department of Biomedical Science and Engineering, Gwangju Institute of Science and Technology, 123 Cheomdangwagi-ro, Buk-gu, Gwangju 61005, Korea E-mail: cmoh@gist.ac.kr
- *Donghyun Kim and Wooju Jeong contributed equally to this study as first authors.
Copyright © 2023 Korean Diabetes Association
This is an Open Access article distributed under the terms of the Creative Commons Attribution Non-Commercial License (http://creativecommons.org/licenses/by-nc/4.0/) which permits unrestricted non-commercial use, distribution, and reproduction in any medium, provided the original work is properly cited.
ABSTRACT
-
Background
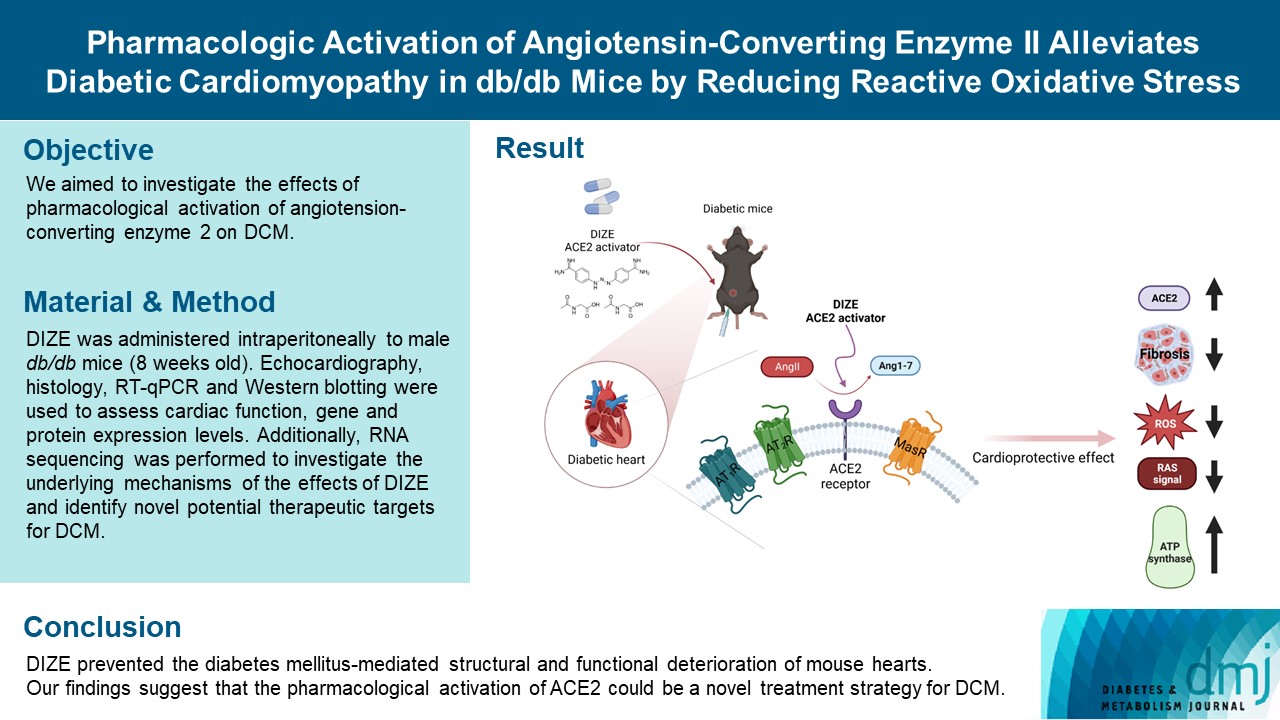
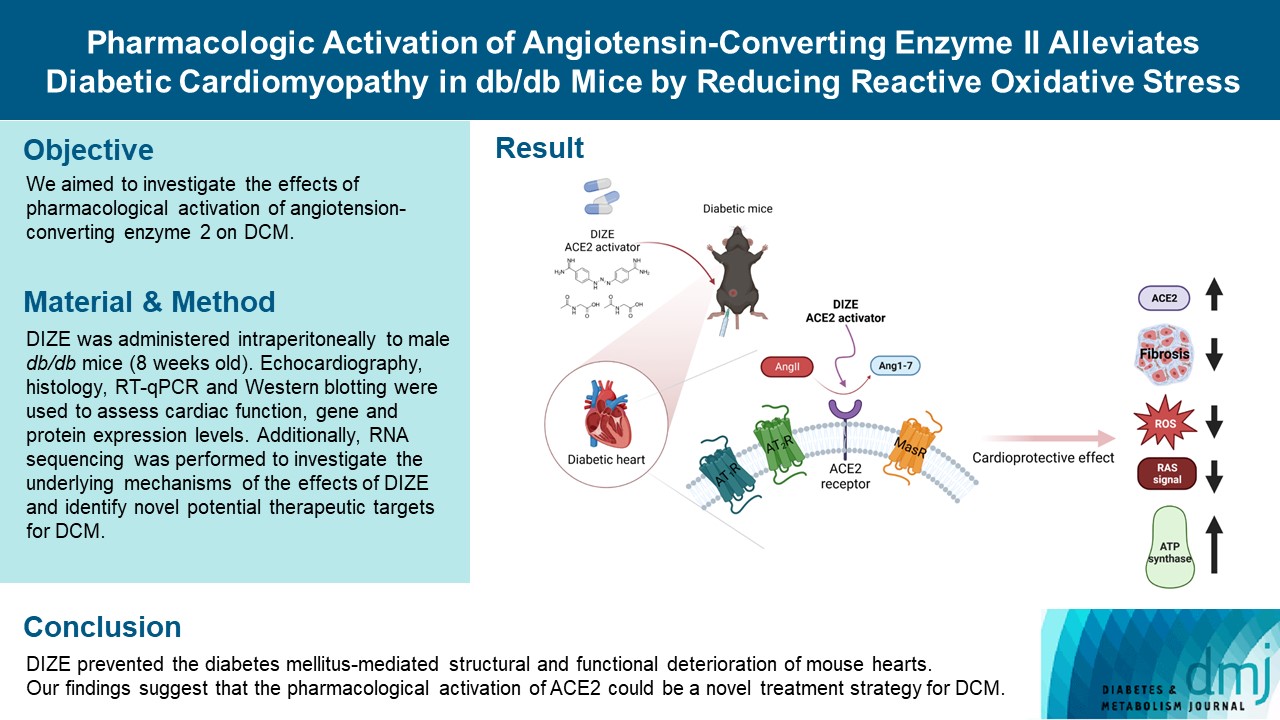
- Diabetes mellitus is one of the most common chronic diseases worldwide, and cardiovascular disease is the leading cause of morbidity and mortality in diabetic patients. Diabetic cardiomyopathy (DCM) is a phenomenon characterized by a deterioration in cardiac function and structure, independent of vascular complications. Among many possible causes, the renin-angiotensin-aldosterone system and angiotensin II have been proposed as major drivers of DCM development. In the current study, we aimed to investigate the effects of pharmacological activation of angiotensin-converting enzyme 2 (ACE2) on DCM.
-
Methods
- The ACE2 activator diminazene aceturate (DIZE) was administered intraperitoneally to male db/db mice (8 weeks old) for 8 weeks. Transthoracic echocardiography was used to assess cardiac mass and function in mice. Cardiac structure and fibrotic changes were examined using histology and immunohistochemistry. Gene and protein expression levels were examined using quantitative reverse transcription polymerase chain reaction and Western blotting, respectively. Additionally, RNA sequencing was performed to investigate the underlying mechanisms of the effects of DIZE and identify novel potential therapeutic targets for DCM.
-
Results
- Echocardiography revealed that in DCM, the administration of DIZE significantly improved cardiac function as well as reduced cardiac hypertrophy and fibrosis. Transcriptome analysis revealed that DIZE treatment suppresses oxidative stress and several pathways related to cardiac hypertrophy.
-
Conclusion
- DIZE prevented the diabetes mellitus-mediated structural and functional deterioration of mouse hearts. Our findings suggest that the pharmacological activation of ACE2 could be a novel treatment strategy for DCM.
- The occurrence of diabetes mellitus (DM) is still on the rise worldwide and is accompanied by a major financial burden [1]. DM and cardiovascular disease (CVD) have a close relationship [2], and the clinical outcomes associated with heart failure (HF) are significantly worse in patients with DM than in those without [3].
- Diabetic cardiomyopathy (DCM) is characterized by the deterioration of cardiac function and structure in the absence of other cardiac risk factors, such as hypertension, coronary atherosclerosis, and valvular heart disease [3]. Major clinical abnormalities in DM, including hyperglycemia, insulin resistance, and hyperinsulinemia, are involved in the development of DCM [3]. Lipotoxicity, caused by circulating free fatty acids and disrupted calcium homeostasis in cardiomyocytes, is a well-known risk factor for DCM pathogenesis [4].
- The renin-angiotensin-aldosterone system (RAAS) has long been proposed as a major contributor to the development of DCM [4]. DM activates RAAS and increases the levels of angiotensin II (Ang II). Hyperglycemia-induced Ang II activation stimulates cardiac fibroblast proliferation and changes in collagen metabolism [5]. Furthermore, elevated blood pressure, sympathetic nerve stimulation, and renal insufficiency caused by Ang II are all remote remodeling processes in the heart [6].
- The activity of the RAAS is counterbalanced by the classical axis—angiotensin-converting enzyme (ACE)/Ang II/Ang II type 1 (AT1) receptor—and the non-classical axis—ACE2/Ang-(1-7)/Mas receptor [7]. The classical RAAS axis stimulates the AT1 receptor and contributes significantly to the pathophysiology of HF by inducing cardiac fibrosis, inflammation, hypertrophy, and cardiac remodeling [8]. The non-classical axis has the opposite effect to the classical axis via an ACE2 dependent pathway [8]. ACE2 modulates cardioprotective activity by reducing deleterious Ang II signaling and generating the protective peptide Ang-(1-7) [9,10]. The most important activity of Ang-(1-7) is vasodilation as well as anti-proliferative and anti-inflammatory effects mediated by Mas receptors [8]. Recently, several studies have reported that depletion of ACE2 worsens DCM, whereas ACE2 activation attenuates Ang II-induced cardiac dysfunction [11,12]. Ang-(1-7) administration also prevents cardiac dysfunction in db/db mice [13]. These findings imply that the modulation of ACE2 activity could be a promising therapeutic strategy for DCM.
- Diminazene aceturate (DIZE) is a small molecule that is approved by the U.S. Food and Drug Administration (FDA) and has been used to treat human trypanosomiasis [14]. It has recently been proposed that DIZE activates ACE2, resulting in increased Ang-(1-7) generation from Ang II [14]. In the present study, we aimed to evaluate the hypothesis that the enhancement of ACE2, using an FDA-approved ACE2 activator (DIZE), would have cardioprotective roles in a murine DCM model.
INTRODUCTION
- Animals
- Male C57BLKS/J-Lepr (db/db) and C57BLKS/J-m+Lepr (db/+) mice were purchased from Jackson Laboratories (Shizuoka Laboratory Center, Shizuoka, Japan). Mice were housed in a 12 hours light-dark cycle at ambient temperature (22°C±1°C) and received a standard chow diet (#2018SC, ENVIGO, North America) and water ad libitum. All tests were performed during the daytime (AM 7:00 to PM 6:00). All experiments were reviewed and approved by Institutional Animal Care and Use Committee of Gwangju Institute of Science and Technology (Approval number: GIST-2021-110).
- Experimental groups and treatment protocol
- We divided 8-week-old male mice into four groups: db/+with vehicle (CTL_VEH), db/+ with DIZE (CTL_DIZE), db/db with vehicle (DB_VEH), and db/db with DIZE (DB_DIZE). Before drug administration, we investigated the left ventricular (LV) systolic function of the mice using echocardiography. DIZE was purchased from Sigma-Aldrich (CAS 908-54-3, St. Louis, MO, USA) and freshly prepared in 0.9% sterile saline at a concentration of 50 mg/mL, once a week. DIZE was administered to mice in the CTL_DIZE and DB_DIZE groups at 15 mg/kg/day by intraperitoneal injection using a BD Ultra-Fine II 8 mm (31G) 1 mL insulin syringe. Mice in the CTL_VEH and DB_VEH groups received equivalent volumes of vehicle (0.9% normal saline) injections. All mice were administered DIZE or saline for 8 weeks. The dosage of DIZE for our study was based on prior preclinical studies [15-18].
- Cellular reactive oxygen species measurement by DCFDA assay
- H9c2, a rat cardiomyoblast cell line, was obtained from a Korean cell line bank and cultured in high-glucose Dulbecco’s Modified Eagle’s medium (DMEM, Invitrogen, Carlsbad, CA, USA) with 10% (V/V) fetal bovine serum (Hyclone, Logan, UT, USA), and 1% (V/V) antibiotic mixture (#15240-062, Gibco, Waltham, MA, USA) at 37°C in a 5% CO2 atmosphere. H2-dichlorodihydrofluorescein diacetate (H2DCFDA, #D399, Invitrogen) was used to assess the intracellular amount of reactive oxygen species (ROS) in the H9c2 cell line. Briefly, for the vehicle and experimental groups, cells were seeded onto 6-well plates and cultured for 24 hours with just media or 100 nM Ang II in medium. Cells in the treatment group were pretreated for 1 hour in medium with 5 μM DIZE before being removed. In addition, 100 nM AngII in medium was incubated for 24 hours. Following the removal of the 2ʹ-7ʹ dichlorofluorescin diacetate (DCFH-DA) and additional washing, the fluorescence of the cells in each well was captured using an upright confocal microscope at 20× with an excitation wavelength of 485 nm and an emission wavelength of 535 nm (BX61WI body, FV1000 model system, Olympus Co. Ltd., Tokyo, Japan).
- Echocardiography
- Mice were anesthetized by inhaled isoflurane (3 L/min), and their cardiac function was assessed using the ACUSON NX3 Elite Ultrasound system (SIEMENS Healthineers, Munich, Germany) with a VF16-5 transducer (16.0 MHz). The LV dimensions, left ventricular ejection fraction (LVEF), left ventricular fractional shortening (LVFS), LV mass, and LV wall thickness were measured.
- Western blot analysis
- Western blotting was performed as previously described [19]. LV tissue samples were disrupted and homogenized using a bead mill homogenizer. Total protein concentration was measured using a DC protein assay kit (Bio-Rad, Hercules, CA, USA). Proteins were separated by electrophoresis and transferred to nitrocellulose membranes. Membranes were blocked in Tris-buffered saline (TBS) containing 0.05% Tween 20 and 5% non-fat dry milk. The membranes were then incubated with specific primary antibodies (1:1,000) recognizing superoxide dismutase 1/2 (Sod1/2; #67480, #66474, Proteintech, Rosemont, IL, USA), and alpha-tubulin (#2144, Cell Signaling Technology, Danvers, MA, USA). After washing with TBS containing 0.05% Tween 20, the membranes were incubated with horseradish peroxidase-linked secondary antibodies (Cell Signaling Technology) and visualized using an ECL detection kit (Sigma-Aldrich; Merck Millipore, Burlington, MA, USA). Signals were captured using luminography, and densitometric analysis was performed using ImageJ software version 1.53 (https://imagej.nih.gov).
- Histological analysis
- Mouse hearts were harvested, fixed in 4% paraformaldehyde (#HP2031, Biosesang, Seongnam, Korea), dehydrated, and embedded in paraffin as previously described [14]. Then, 4 µm-thick tissue sections were deparaffinized and stained with hematoxylin and eosin (H&E). Fibrosis was detected by staining with Picrosirius red, according to the manufacturer’s protocol. ImageJ was used to quantify the fibrotic area, and each slide was assessed by an independent pathologist in a blinded manner for confirmation.
- Blood biochemistry
- Serum was prepared from the blood by incubating at room temperature for 20 minutes and subsequent centrifugation at 2,500 rpm for 20 minutes at 4°C. Serum levels of Ang-(1-7) were measured using the mouse Ang-(1-7) RTU ELISA kit (#MBS457587, MyBioSource, Vancouver, BC, Canada), according to the manufacturer’s instructions. Plasma was prepared from the blood by centrifugation at 2,000 ×g for 20 minutes at 4°C. Plasma levels of Ang II were measured using a mouse Ang II ELISA kit (#MBS022429, MyBioSource), according to the manufacturer’s instructions.
- Myocardial gene expression analysis
- Frozen LV samples were disrupted and homogenized using a bead mill homogenizer. RNA extraction and quantitative reverse transcription polymerase chain reaction (qRT-PCR) were performed. Total RNA was extracted using TRIzol reagent (Invitrogen, Thermo Fisher Scientific, Waltham, MA, USA) according to the manufacturer’s protocol. Total RNA (1 µg) was reverse-transcribed using the High-Capacity cDNA Reverse Transcription kit (Applied Biosystems, Waltham, MA, USA; Thermo Fisher Scientific) according to the manufacturer’s protocol. qRT-PCR was performed using amfiSure qGreen Q-PCR master mix (#Q5602, GenDEPOT, Katy, TX, USA). Primer sequences are provided in Supplementary Table 1.
- Identification of differentially expressed genes
- Raw data were processed using the ‘edgeR’ package [20] with R software version 4.1 (R Foundation for Statistical Computing, Vienna, Austria) to generate counts per million (cpm), which were then transformed to a log2 scale and standardized using quantile normalization. The adjusted log2-cpm was then used in an integrative statistical technique to detect the differentially expressed genes (DEGs). For each gene, the Student’s t-test and log2-median-ratio were used to calculate the observed T-value and the log2-median-ratio between the two conditions. Subsequently, random sampling of 1,000 times was used to produce empirical null distributions for the T-value and log2-median-ratio.
- To obtain an overall P value for each gene, we first computed the adjusted P values by running a two-tailed test on the measured T-value and log2-median-ratio using their respective empirical distributions. The adjusted P values were then merged using Stouffer’s method. DEGs were chosen for each comparison based on two criteria: an overall P value of less than 0.05, and an absolute log2-median-ratio greater than the median of the empirical distribution’s 2.5th and 97.5th percentiles for the log2-median-ratio.
- Pathway analysis: KEGG, GO analysis, and GSEA
- Functional enrichment analysis of the DEGs was performed using Database for Annotation, Visualization and Integrated Discovery (DAVID) software (https://david.ncifcrf.gov) [21]. Our Gene Ontology (GO) biological processes (BPs) and Kyoto Encyclopedia of Genes and Genomes (KEGGs) were selected using two criteria: a P value of less than 0.05 and a gene count greater than three. For visualization in the heat map, we converted the enrichment P value into a Z-score by Z=N−1(1-P); where N−1(∙) is the inverse standard normal distribution. Gene Set Enrichment Analysis (GSEA) was used to assess the enrichment of DEGs using the ‘clusterProfiler’ package [22] in R software version 4.1. We conducted the GSEA analysis with 10,000 permutations, 3 minGSSize, 800 maxGSSize, and pAdjustMethod “BH”.
- Cellular process network analysis
- We built a process network for GO BPs and KEGG pathways enriched by upregulated and downregulated terms, each with at least five upregulated and downregulated genes. A list of genes annotated with the corresponding GO BP/KEGG pathways for each node was obtained. Every node is clustered through a community cluster (GLAY) in ‘clusterMaker’ [23]. If the number of common genes was greater than three, then each pair of nodes was connected to an edge. Each edge connecting nodes i and j was then quantified by measuring the Sørensen-Dice coefficient, as previously described [24,25], which is defined as 2|Pi ∩Pj|/(|Pi|+ |Pj|); where |Pi|and |Pj|are the numbers of genes in nodes i and j and |Pi ∩Pj|is the number of common genes. Finally, the process network was visualized using Cytoscape software (https://cytoscape.org) [26] in the organic layout mode, where only the edges with Sørensen-Dice coefficients >0.3673 were included. A cutoff of 0.3673 was calculated using the 90th percentile of the coefficient distribution of all possible pairs of nodes in the network.
- Upstream regulator analysis
- The commercial QIAGEN Ingenuity Pathway Analysis software (IPA, QIAGEN Redwood City, www.qiagen.com/ingenuity) was used to perform an upstream regulatory analysis (URA) of the DEGs discovered in our analysis [27]. The P value was computed using Fisher’s exact test, with a significance cut-off of 0.05.
- Statistical analysis
- All statistical analyses were performed using GraphPad Prism 9.0.0 (GraphPad Software Inc., San Diego, CA, USA). Differences between groups were evaluated for statistical significance using the Student’s t-test or one-way analysis of variance (ANOVA), followed by Tukey’s post hoc test and Sidak’s multiple-comparison test. Statistical significance was set at P<0.05.
METHODS
- DIZE attenuates DCM in db/db mice
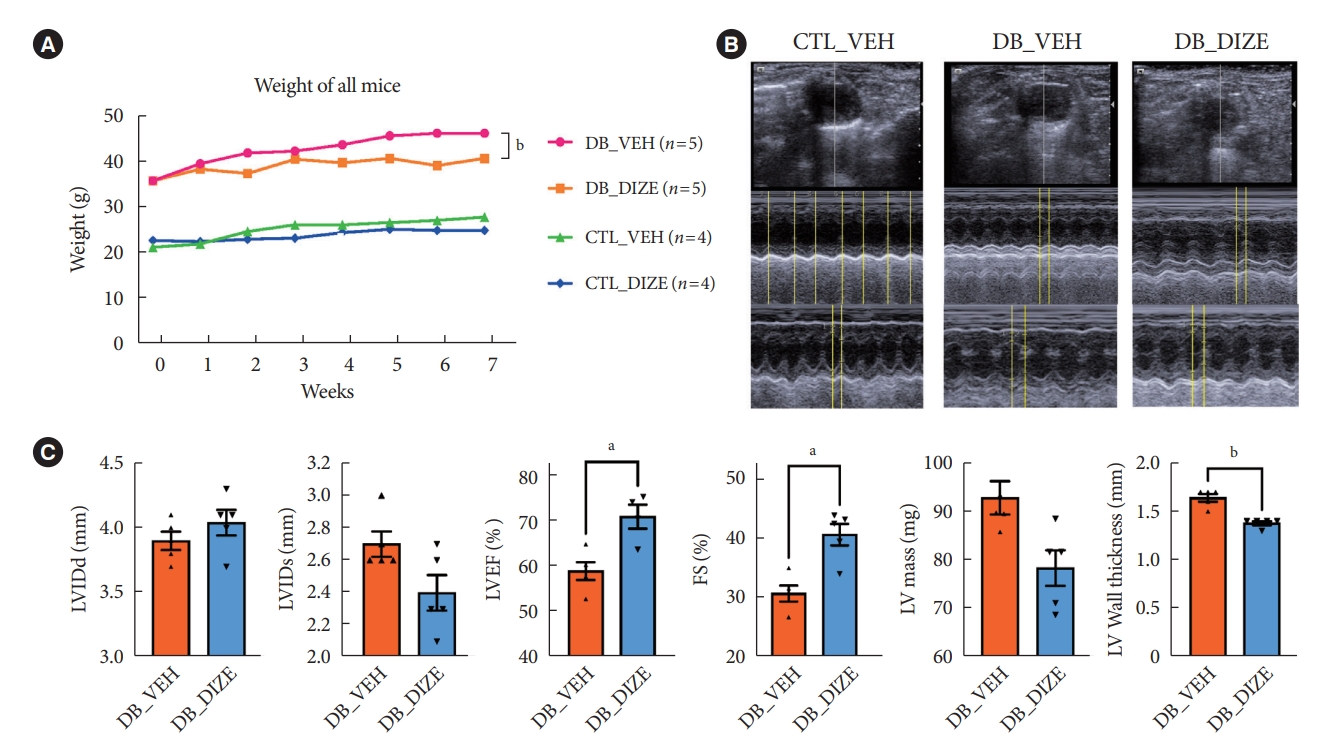
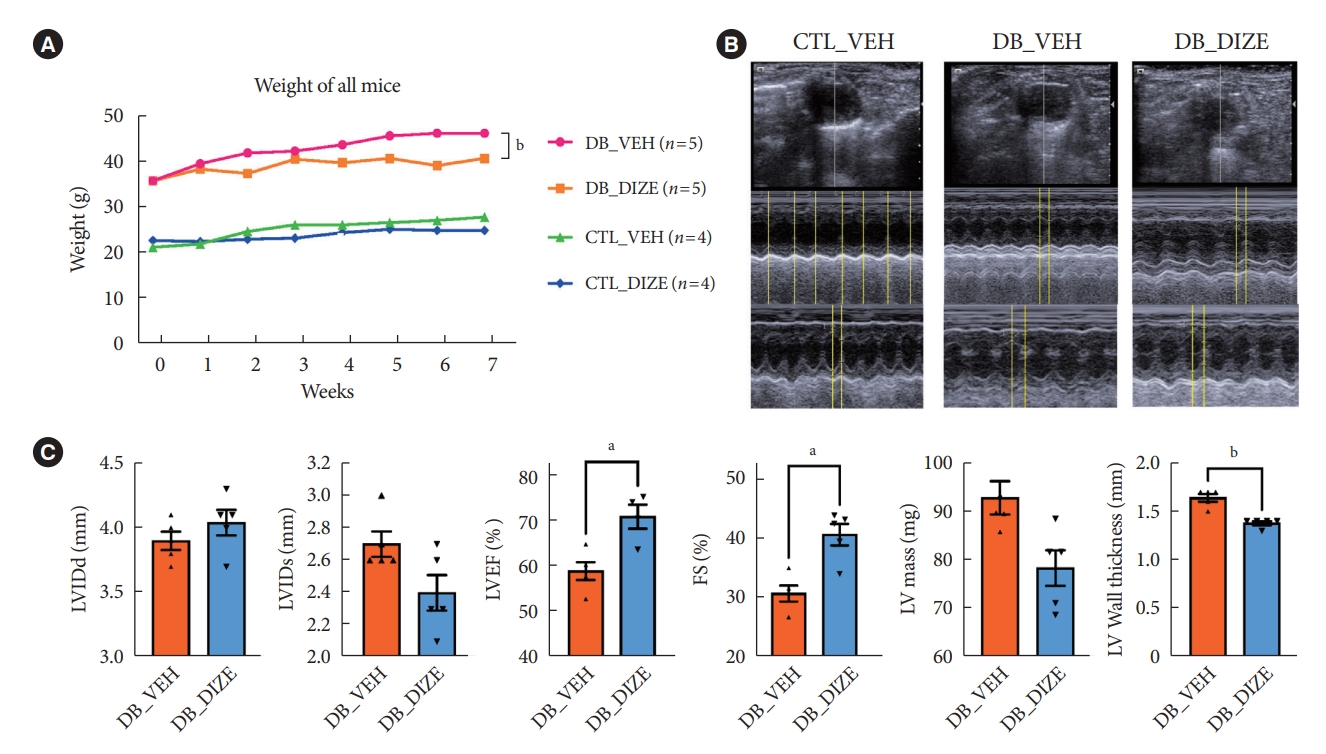
- To investigate the beneficial effects of ACE2 activation on diabetic hearts, we administered DIZE to db/+ and db/db mice for 8 weeks. During this period, the bodyweight changes were significantly different between the vehicle- and DIZE-treated mice (Fig. 1A), where the vehicle-treated db/db mice gained more weight than the DIZE-treated db/db mice. Echocardiography was performed to evaluate whether DIZE prevents DCM in db/db mice (Fig. 1B). The DB_DIZE group had significantly higher LVEF and LVFS as well as lower LV wall thickness than the DB_VEH group; this indicates improved systolic function and less cardiac hypertrophy (Fig. 1C).
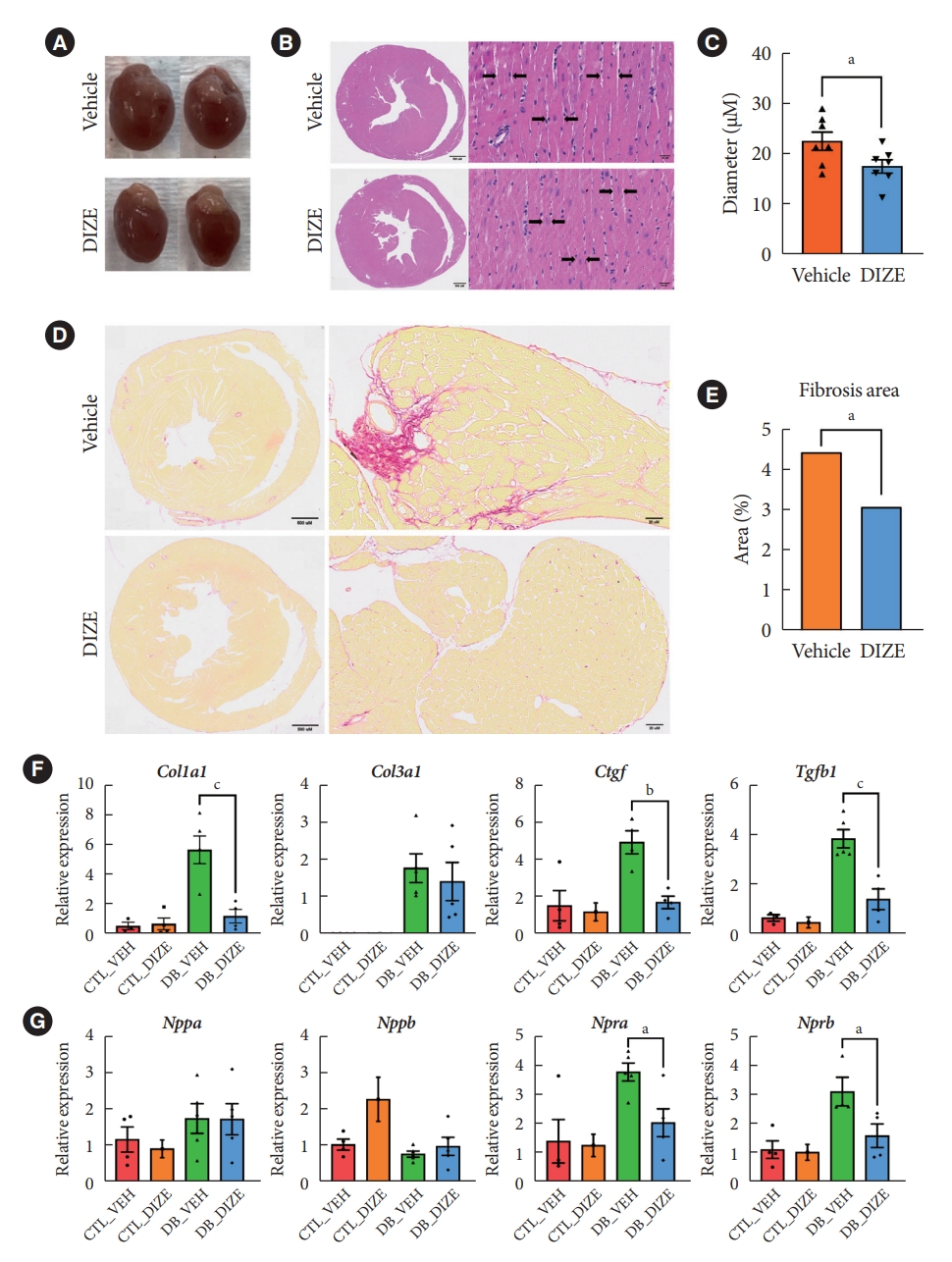
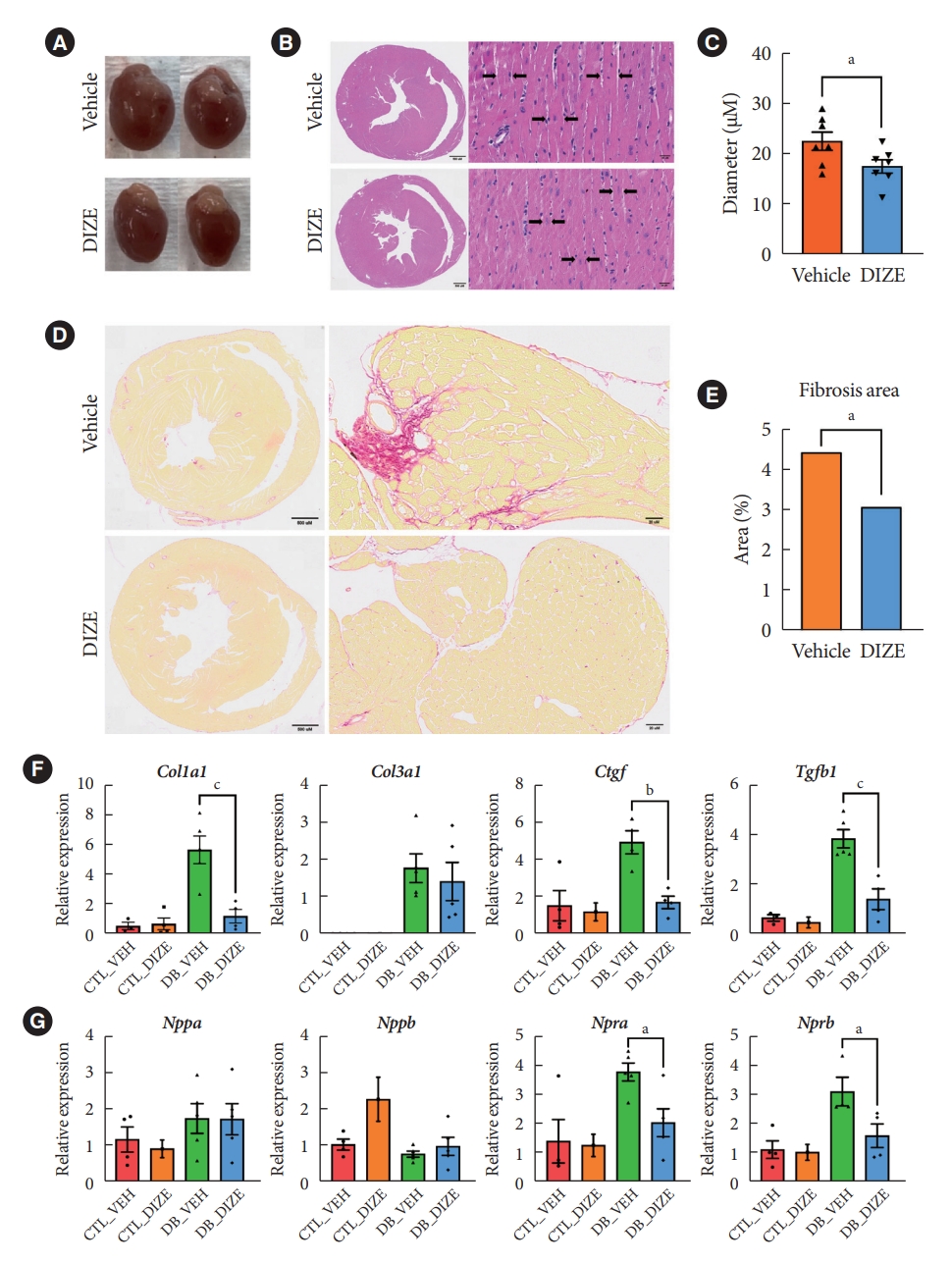
- DIZE reduces cardiac fibrosis in db/db mice
- Next, we performed histological analyses to confirm the cardiac benefits of DIZE treatment in DCM hearts (Fig. 2). Echocardiography (Fig. 1C) and a gross view (Fig. 2A) show that DIZE treatment tended to decrease heart size in db/db mice; however, the difference was not significant. Although, myofibril diameters were significantly decreased in DB_DIZE hearts compared with DB_VEH hearts (Fig. 2B and C). Picrosirius red staining revealed that DIZE reduced perivascular and interstitial fibrosis in the hearts of db/db mice (Fig. 2D and E). In line with these findings, fibrosis-related gene expression, such as the collagen type I alpha 1 chain (Col1a1), connective tissue growth factor (Ctgf), and transforming growth factor-beta 1 (Tgfβ1) genes, were significantly decreased in the DB_DIZE hearts compared to those of the DB_VEH group (Fig. 2F). Natriuretic peptide hormones are markers of cardiac hypertrophy and HF [28]. Although the expression of natriuretic peptide A (Nppa) and natriuretic peptide B (Nppb) genes did not differ between DB_VEH and DB_DIZE hearts, the expression of atrial natriuretic peptide receptor type A (Npra) gene was significantly lower in DB_DIZE than that in DB_VEH (Fig. 2G).
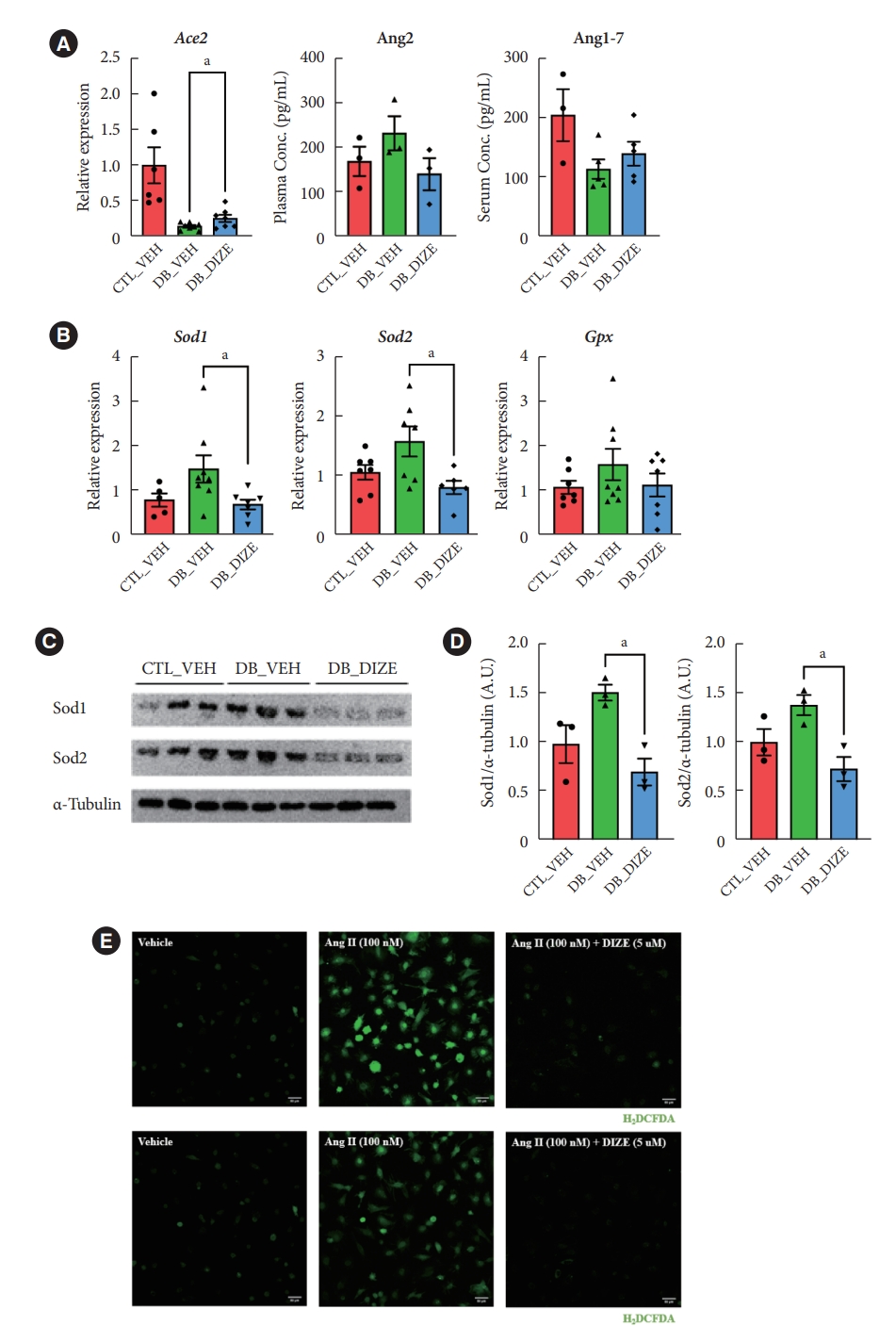
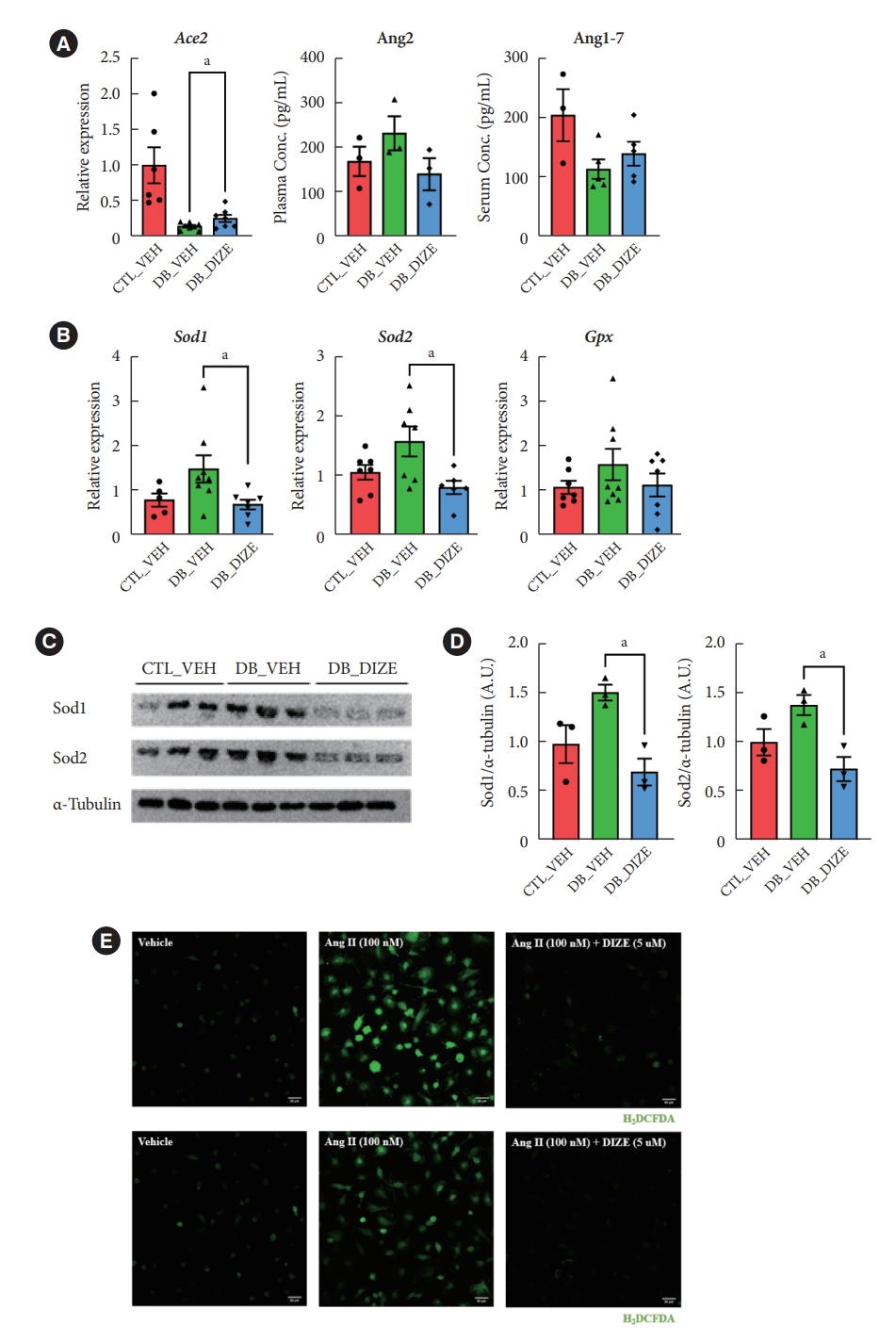
- DIZE increases Ace2 gene expression and reduces oxidative stress in db/db mice
- Previously, several studies found that DIZE increased Ace2 gene expression in major organs, such as the lung, liver, and especially in the ischemic heart, while also increasing ACE2 activity [29]. Therefore, we investigated the effects of DIZE on Ace2 expression in diabetic hearts. As expected, the expression level of Ace2 was significantly decreased in DB_VEH hearts compared to the CTL_VEH group; additionally, the expression was upregulated in the DB_DIZE hearts compared to that in the DB_VEH ones (Fig. 3A). When we compared DB_DIZE blood to DB_VEH blood, the circulating Ang II levels showed a trend toward downregulation, while the Ang-(1-7) levels exhibited a trend toward upregulation (Fig. 3A). However, these differences were not statistically significant.
- Oxidative stress caused by hyperglycemia, hyperlipidemia, hypertension, and inflammation in the diabetic myocardium is a major risk factor for DCM [30]. DIZE significantly reduced the expression of oxidative stress-related genes in the hearts of db/db mice (Fig. 3B). Our Western blot assay produced results that were consistent with those of qRT-PCR (Fig. 3C and D). To evaluate the direct effect of DIZE on Ang II-stimulated ROS, we used H9C2 cardiomyocytes in an in vitro DCFDA experiment. ROS were successfully decreased by DIZE therapy (Fig. 3E). These results demonstrate that ACE2 activators reduce oxidative stress and, thus, prevent myocardial dysfunction in diabetic hearts.
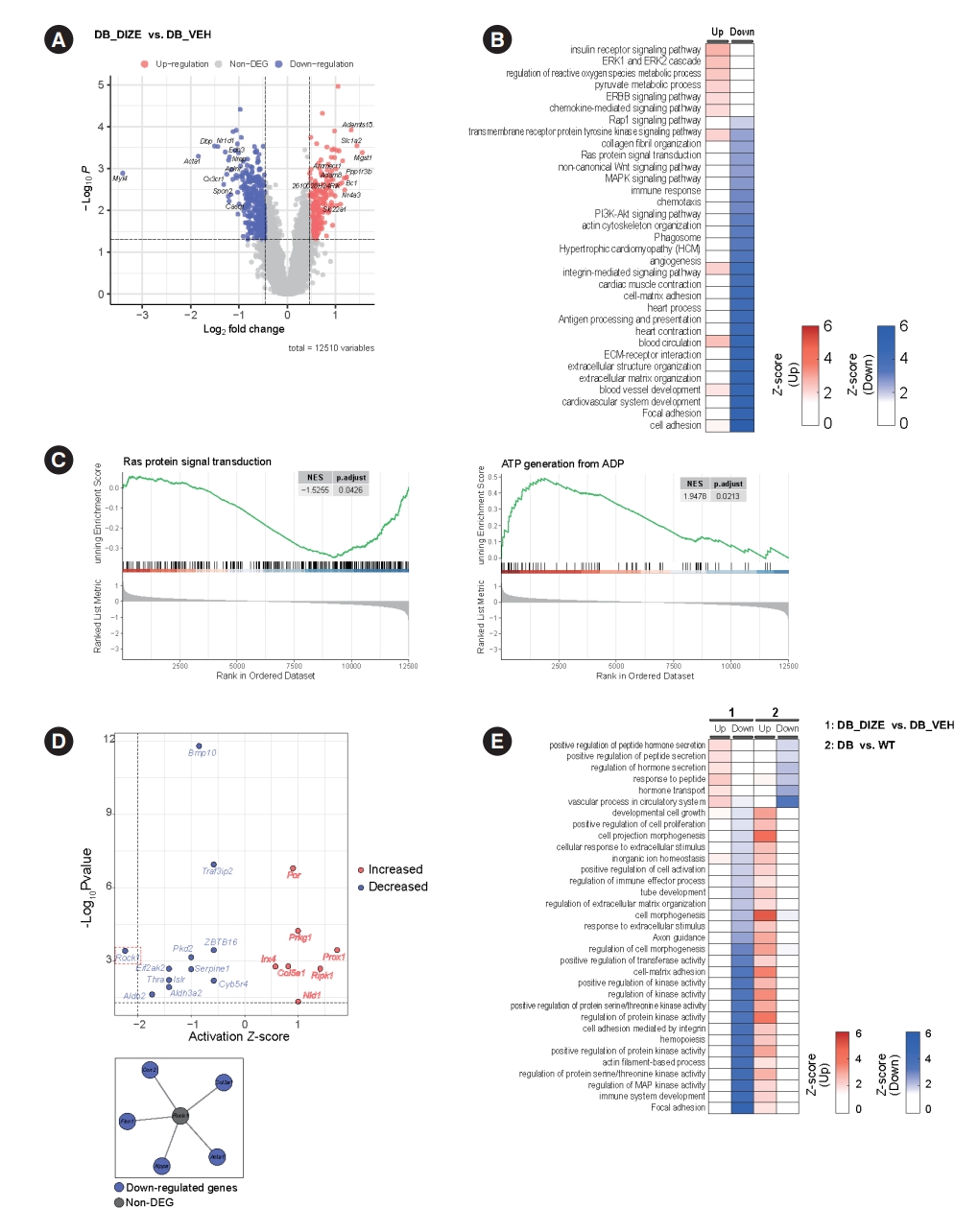
- Transcriptome analysis of the beneficial effect of ACE2 activation
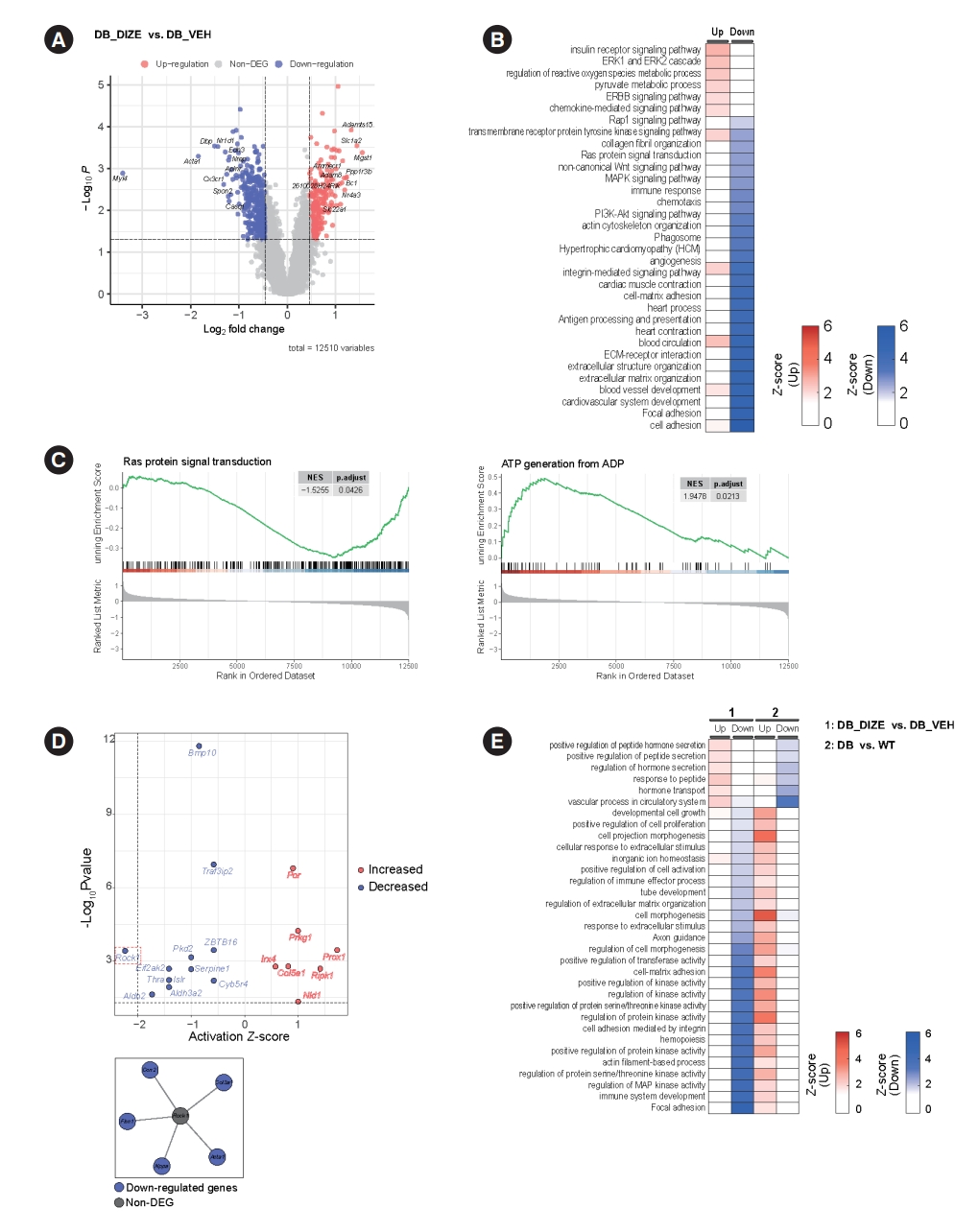
- Our next objective was to identify possible molecular mechanisms underlying the beneficial effects of DIZE on DCM. Therefore, we performed RNA sequencing and profiled the heart transcriptomes of the DB_VEH and DB_DIZE groups. Fig. 4A shows the volcano plot, while the DEGs with satisfying values (overall P<0.05, log2 fold change ≥0.462460853, and log2 fold change ≤–0.462460853; with 2.5th and 97.5th percentiles of the empirical distribution for the log2-median ratio) are labeled in 234 red dots (upregulated DEGs) and 394 blue dots (downregulated DEGs). The top five DEGs were microsomal glutathione transferase 1 (MGST1), solute carrier family 1 member 2 (Slc1a2), ADAM metallopeptidase with thrombospondin type 1 motif 15 (Adamts15), protein phosphatase 1 regulatory subunit 3 B (Ppp1r3b), and brain cytoplasmic RNA 1 (Bc1). The most common DEGs were myosin light chain 4 (Myl4), actin alpha 1, skeletal muscle (Acta1), DNA-binding protein phosphatase 1 (Dbp1), nuclear receptor subfamily 1 group D member 1 (Nr1d1), and C-X3-C motif chemokine receptor 1 (Cx3cr1).
- To study the functional enrichment of DEGs, we performed GO and KEGG pathway analyses. Intriguingly, DIZE upregulates several pathways related to insulin receptor signaling, the extracellular signal-regulated kinase 1/2 (ERK1/2) cascade, and the process of angiogenesis. DIZE also downregulates several pathways related to collagen fibril organization, immune response, and the extracellular matrix (Fig. 4B). These pathways are important components of DCM pathophysiology. GSEA enrichment plots showed that DIZE administration was negatively associated with Ras protein signaling, which is a major therapeutic target in cardiac hypertrophy and HF (Fig. 4C) [31]. Furthermore, DIZE showed positive associations with adenosine triphosphate (ATP) generation, which is a major problem in HF (Fig. 4C) [32]. Based on the enriched pathways, we performed a cellular process network analysis using Cytoscape software. The analysis revealed that the most highly connected cellular networks were related to ‘antigen processing and presentation’, ‘angiogenesis’, and ‘cardiac muscle contraction’ (Supplementary Fig. 1).
- To identify potential upstream molecules, we performed pathway and URAs using QIAGEN’s IPA tool. The results are shown in Fig. 4D. The top upstream activated regulators were predicted to be the prospero homeobox protein 1 (Prox1) and receptor-interacting serine/threonine kinase 1 (Ripk1) genes. The top downregulated regulators were the Rho-associated coiled-coil containing protein kinase 1 (Rock1) and the aldehyde dehydrogenase 2 family member (Aldh2) genes.
- Next, we compared the enriched pathways related to DCM pathophysiology with those related to DIZE administration. For this, we used the publicly available transcriptome profiling dataset from the NCBI GEO database (GSE161931) [33]. GSE161931 is a cardiac RNA-sequencing dataset obtained from db/db and C57BLKS/J (BKS) mice. We identified DEGs in the heart between the DCM and control groups (Supplementary Fig. 2) and performed pathway analysis based on these DEGs. Fig. 4E shows the enriched pathways from our original transcriptome data and the public transcriptome data, which pointed in opposite directions. Diabetic hearts showed upregulated pathways related to ‘cell morphogenesis,’ ‘cell-matrix adhesion,’ and the ‘regulation of protein kinase activity.’ These pathways were downregulated by DIZE treatment (Fig. 4E). ‘Vascular process in circulatory system,’ ‘regulation of hormone secretion,’ and ‘hormone transport’ were found to be downregulated in the diabetic hearts compared to controls, although DIZE treatment upregulated these pathways (Fig. 4E).
RESULTS
- In the current study, we investigated the effects of ACE2 activators on cardiac dysfunction in the diabetic hearts of db/db mice. DIZE, an ACE2 activator, did not affect blood glucose levels but significantly prevented DCM progression in the hearts of db/db mice (Supplementary Fig. 3). We found that DIZE administration for 8 weeks improved systolic function and reduced cardiac fibrosis.
- During this period, DIZE also reduced weight gain in db/db mice (Fig. 1A). Although there is a limit to the inability to measure food and water intake, this was consistent with previous findings that Ang-(1-7) reduced fat mass and improved adipose tissue inflammation in db/db mice and high-fat dietinduced obese mice [13,34]. Also, administration of recombinant human ACE2 (rhACE2) to high-fat diet-induced obese mice showed similar results. rhACE2 increases oxygen consumption and upregulates thermogenesis in an obese mouse model by activating brown fat metabolism [35]. In our study, there are limitations that the weight of each organ could not be measure while focusing on cardiac function, but the results observed by Morimoto et al. [34] could represent Ang-(1-7) treatment enlarges the volume of brown adipose tissue, resulting in improved obesity. The bodyweight difference between the DB_VEH and DB_DIZE groups in our study also supports these previous findings.
- Several studies have described the effects of DIZE on tissue fibrosis. DIZE reduced cardiac fibrosis and diastolic dysfunction [36]. DIZE improves liver fibrosis in mice by inhibiting the activation of fibroblastic stellate cells [29]. We also found that DIZE treatment reduced oxidative stress in db/db mouse hearts. The expression of oxidative stress-related genes and proteins decreased (Fig. 3B and C). Pathway analysis also showed that DIZE caused significant changes related to the regulation of reactive oxygen species’ metabolic processes (Fig. 4B). Indeed, previous studies have found that Ang II is a prooxidant peptide [37], and myocardial injury by oxidative stress plays a critical role in the development of DCM [38]. These findings led us to speculate that reduced oxidative stress is the underlying mechanism for the beneficial effects of DIZE in diabetic hearts.
- Transcriptome analysis identified possible targets of DCM and revealed the potential mechanisms of DIZE effects on diabetic hearts. Mgst1 is the most highly upregulated DEG in the DB_DIZE group (Fig. 4A) and it encodes a protein called MGST1, which belongs to the membrane-associated proteins in eicosanoid and glutathione metabolism family [39]. This protein is well known for its role in the conjugation of electrophiles, protection from oxidative stress, and inhibition of ferroptosis [39,40]. Oxidative stress is a major risk factor for DCM. Furthermore, suppression of ferroptosis has been recognized as a novel therapeutic target for CVD [41]. These findings imply that increasing Mgst1 expression using DIZE or other methods could be a therapeutic strategy for DCM.
- Among the predicted upstream regulators, Rock1 was the most suppressed upstream regulator. This gene encodes a serine/threonine kinase and is a downstream target of the small GTPase Rho [42]. Previous studies have shown that ROCK signaling plays various roles in the pathogenesis of CVDs [42]. ROCK1 signaling is involved in pressure overload-induced cardiac hypertrophy [43] and perivascular fibrosis caused by Ang II infusion in a mouse model [44]. Several studies using ROCK inhibitors, such as fasudil and Y27632, have also shown the beneficial effects of ROCK inhibition on cardiac remodeling [45-47]. These findings suggest that DIZE administration may induce beneficial cellular and molecular changes, similar to ROCK1 signaling suppression.
- Our study has several limitations. First, we did not rule out the possibility that the beneficial effects were caused by the off-target effects of DIZE. More research is needed to confirm the beneficial effects of other ACE2 activators in a diabetic animal model, such as xantehnone [48]. Second, during DIZE administration, a few mice died during drug treatment, and histological analyses revealed severe liver toxicity (Supplementary Fig. 4). Although DIZE is known to be beneficial in liver disease models [29], this might be one of the severe side effects of long-term DIZE use. Even if the dosage and treatment duration of DIZE for our study was based on prior preclinical studies [15-18], the inability to proceed the experiments on dose-dependent action of DIZE would be another limitation. Further studies on the toxicity of DIZE are warranted.
- In conclusion, DIZE prevented structural and functional deterioration of the mouse heart caused by diabetes. To the best of our knowledge, this is the first study to report the effects of pharmacological ACE2 activators in DCM. Based on our data, the pharmacological activation of ACE2 may be a novel treatment strategy for DCM.
DISCUSSION
SUPPLEMENTARY MATERIALS
Supplementary Table 1.
Supplementary Fig. 1.
Supplementary Fig. 2.
Supplementary Fig. 3.
Supplementary Fig. 4.
-
CONFLICTS OF INTEREST
No potential conflict of interest relevant to this article was reported.
-
AUTHOR CONTRIBUTIONS
Conception or design: S.W.C., C.M.O., R.P.
Acquisition, analysis, or interpretation of data: D.K., W.J., Y.K., J.L.
Drafting the work or revising: D.K., W.J., Y.K., S.W.C., C.M.O., R.P.
Final approval of the manuscript: D.K., W.J., Y.K., J.L., S.W.C., C.M.O., R.P.
-
FUNDING
This research was supported by the Basic Science Research Program through the National Research Foundation of Korea (NRF), funded by the Korean government (2018R1A5-A1024-340 to Raekil Park), and the Ministry of Education (2020R1C1C1004999 to Chang-Myung Oh, 2020R1C1-C1015104 to Sung Woo Cho), and by the Korea Health Industry Development Institute (KHIDI)-AZ Diabetes Research Program to Chang-Myung Oh, and Samjin pharm Co. to Sung Woo Cho, and the MD-PhD/Medical Scientist Training Program through the KHIDI, funded by the Ministry of Health & Welfare to Donghyun Kim. The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
NOTES
-
Acknowledgements
- None
- 1. Cho NH, Shaw JE, Karuranga S, Huang Y, da Rocha Fernandes JD, Ohlrogge AW, et al. IDF Diabetes Atlas: global estimates of diabetes prevalence for 2017 and projections for 2045. Diabetes Res Clin Pract 2018;138:271-81.ArticlePubMed
- 2. Raghavan S, Vassy JL, Ho YL, Song RJ, Gagnon DR, Cho K, et al. Diabetes mellitus-related all-cause and cardiovascular mortality in a national cohort of adults. J Am Heart Assoc 2019;8:e011295.ArticlePubMedPMC
- 3. Jia G, Hill MA, Sowers JR. Diabetic cardiomyopathy: an update of mechanisms contributing to this clinical entity. Circ Res 2018;122:624-38.ArticlePubMedPMC
- 4. Sun L, Yu M, Zhou T, Zhang S, He G, Wang G, et al. Current advances in the study of diabetic cardiomyopathy: from clinicopathological features to molecular therapeutics (Review). Mol Med Rep 2019;20:2051-62.ArticlePubMed
- 5. Fiordaliso F, Leri A, Cesselli D, Limana F, Safai B, Nadal-Ginard B, et al. Hyperglycemia activates p53 and p53-regulated genes leading to myocyte cell death. Diabetes 2001;50:2363-75.ArticlePubMedPDF
- 6. Boudina S, Abel ED. Diabetic cardiomyopathy, causes and effects. Rev Endocr Metab Disord 2010;11:31-9.ArticlePubMedPMCPDF
- 7. Zhang F, Liu J, Li SF, Song JX, Ren JY, Chen H. Angiotensin-(1-7): new perspectives in atherosclerosis treatment. J Geriatr Cardiol 2015;12:676-82.PubMedPMC
- 8. Harvey B, Jaisser F. Aldosterone-mineralocorticoid receptor: cell biology to translational medicine. IntechOpen; 2019 Chapter 5, Renin-angiotensin-aldosterone system in heart failure: focus on nonclassical angiotensin pathways as novel upstream targets regulating aldosterone [cited 2022 Jul 12]. Available from:https://doi.org/10.5772/intechopen.87239.Article
- 9. Der Sarkissian S, Grobe JL, Yuan L, Narielwala DR, Walter GA, Katovich MJ, et al. Cardiac overexpression of angiotensin converting enzyme 2 protects the heart from ischemia-induced pathophysiology. Hypertension 2008;51:712-8.ArticlePubMed
- 10. Zhao YX, Yin HQ, Yu QT, Qiao Y, Dai HY, Zhang MX, et al. ACE2 overexpression ameliorates left ventricular remodeling and dysfunction in a rat model of myocardial infarction. Hum Gene Ther 2010;21:1545-54.ArticlePubMed
- 11. Patel VB, Bodiga S, Basu R, Das SK, Wang W, Wang Z, et al. Loss of angiotensin-converting enzyme-2 exacerbates diabetic cardiovascular complications and leads to systolic and vascular dysfunction: a critical role of the angiotensin II/AT1 receptor axis. Circ Res 2012;110:1322-35.ArticlePubMedPMC
- 12. Dong B, Yu QT, Dai HY, Gao YY, Zhou ZL, Zhang L, et al. Angiotensin-converting enzyme-2 overexpression improves left ventricular remodeling and function in a rat model of diabetic cardiomyopathy. J Am Coll Cardiol 2012;59:739-47.ArticlePubMed
- 13. Mori J, Patel VB, Abo Alrob O, Basu R, Altamimi T, Desaulniers J, et al. Angiotensin 1-7 ameliorates diabetic cardiomyopathy and diastolic dysfunction in db/db mice by reducing lipotoxicity and inflammation. Circ Heart Fail 2014;7:327-39.ArticlePubMed
- 14. Qaradakhi T, Gadanec LK, McSweeney KR, Tacey A, Apostolopoulos V, Levinger I, et al. The potential actions of angiotensin-converting enzyme II (ACE2) activator diminazene aceturate (DIZE) in various diseases. Clin Exp Pharmacol Physiol 2020;47:751-8.ArticlePubMedPDF
- 15. Evans CE, Miners JS, Piva G, Willis CL, Heard DM, Kidd EJ, et al. ACE2 activation protects against cognitive decline and reduces amyloid pathology in the Tg2576 mouse model of Alzheimer’s disease. Acta Neuropathol 2020;139:485-502.ArticlePubMedPMCPDF
- 16. Goru SK, Kadakol A, Malek V, Pandey A, Sharma N, Gaikwad AB. Diminazene aceturate prevents nephropathy by increasing glomerular ACE2 and AT2 receptor expression in a rat model of type1 diabetes. Br J Pharmacol 2017;174:3118-30.ArticlePubMedPMCPDF
- 17. Duan R, Xue X, Zhang QQ, Wang SY, Gong PY, Yan E, et al. ACE2 activator diminazene aceturate ameliorates Alzheimer’s disease-like neuropathology and rescues cognitive impairment in SAMP8 mice. Aging (Albany NY) 2020;12:14819-29.ArticlePubMedPMC
- 18. Kamel AS, Abdelkader NF, Abd El-Rahman SS, Emara M, Zaki HF, Khattab MM. Stimulation of ACE2/ANG(1-7)/Mas axis by diminazene ameliorates Alzheimer’s disease in the D-galactose-ovariectomized rat model: role of PI3K/Akt pathway. Mol Neurobiol 2018;55:8188-202.ArticlePubMedPDF
- 19. Oh CM, Cho S, Jang JY, Kim H, Chun S, Choi M, et al. Cardioprotective potential of an SGLT2 inhibitor against doxorubicin-induced heart failure. Korean Circ J 2019;49:1183-95.ArticlePubMedPMCPDF
- 20. Robinson MD, McCarthy DJ, Smyth GK. edgeR: a Bioconductor package for differential expression analysis of digital gene expression data. Bioinformatics 2010;26:139-40.PubMed
- 21. Huang da W, Sherman BT, Lempicki RA. Systematic and integrative analysis of large gene lists using DAVID bioinformatics resources. Nat Protoc 2009;4:44-57.ArticlePubMedPDF
- 22. Subramanian A, Tamayo P, Mootha VK, Mukherjee S, Ebert BL, Gillette MA, et al. Gene set enrichment analysis: a knowledge-based approach for interpreting genome-wide expression profiles. Proc Natl Acad Sci U S A 2005;102:15545-50.ArticlePubMedPMC
- 23. Morris JH, Apeltsin L, Newman AM, Baumbach J, Wittkop T, Su G, et al. clusterMaker: a multi-algorithm clustering plugin for Cytoscape. BMC Bioinformatics 2011;12:436.ArticlePubMedPMCPDF
- 24. Dice LR. Measures of the amount of ecologic association between species. Ecology 1945;26:297-302.ArticlePDF
- 25. Mueller LN, Rinner O, Schmidt A, Letarte S, Bodenmiller B, Brusniak MY, et al. SuperHirn: a novel tool for high resolution LC-MS-based peptide/protein profiling. Proteomics 2007;7:3470-80.ArticlePubMedPDF
- 26. Shannon P, Markiel A, Ozier O, Baliga NS, Wang JT, Ramage D, et al. Cytoscape: a software environment for integrated models of biomolecular interaction networks. Genome Res 2003;13:2498-504.ArticlePubMedPMC
- 27. Kramer A, Green J, Pollard J Jr, Tugendreich S. Causal analysis approaches in ingenuity pathway analysis. Bioinformatics 2014;30:523-30.ArticlePubMedPDF
- 28. Sergeeva IA, Christoffels VM. Regulation of expression of atrial and brain natriuretic peptide, biomarkers for heart development and disease. Biochim Biophys Acta 2013;1832:2403-13.ArticlePubMed
- 29. Rajapaksha IG, Mak KY, Huang P, Burrell LM, Angus PW, Herath CB. The small molecule drug diminazene aceturate inhibits liver injury and biliary fibrosis in mice. Sci Rep 2018;8:10175.ArticlePubMedPMCPDF
- 30. Liu Q, Wang S, Cai L. Diabetic cardiomyopathy and its mechanisms: role of oxidative stress and damage. J Diabetes Investig 2014;5:623-34.ArticlePubMedPMCPDF
- 31. Ramos-Kuri M, Meka SH, Salamanca-Buentello F, Hajjar RJ, Lipskaia L, Chemaly ER. Molecules linked to Ras signaling as therapeutic targets in cardiac pathologies. Biol Res 2021;54:23.ArticlePubMedPMCPDF
- 32. Ingwall JS. Energy metabolism in heart failure and remodelling. Cardiovasc Res 2009;81:412-9.ArticlePubMed
- 33. Huang X, Zhang KJ, Jiang JJ, Jiang SY, Lin JB, Lou YJ. Identification of crucial genes and key functions in type 2 diabetic hearts by bioinformatic analysis. Front Endocrinol (Lausanne) 2022;13:801260.ArticlePubMedPMC
- 34. Morimoto H, Mori J, Nakajima H, Kawabe Y, Tsuma Y, Fukuhara S, et al. Angiotensin 1-7 stimulates brown adipose tissue and reduces diet-induced obesity. Am J Physiol Endocrinol Metab 2018;314:E131-8.ArticlePubMed
- 35. Kawabe Y, Mori J, Morimoto H, Yamaguchi M, Miyagaki S, Ota T, et al. ACE2 exerts anti-obesity effect via stimulating brown adipose tissue and induction of browning in white adipose tissue. Am J Physiol Endocrinol Metab 2019;317:E1140-9.ArticlePubMed
- 36. Velkoska E, Patel SK, Griggs K, Burrell LM. Diminazene aceturate improves cardiac fibrosis and diastolic dysfunction in rats with kidney disease. PLoS One 2016;11:e0161760.ArticlePubMedPMC
- 37. Rabelo LA, Alenina N, Bader M. ACE2-angiotensin-(1-7)-Mas axis and oxidative stress in cardiovascular disease. Hypertens Res 2011;34:154-60.ArticlePubMedPDF
- 38. Cai L, Kang YJ. Oxidative stress and diabetic cardiomyopathy: a brief review. Cardiovasc Toxicol 2001;1:181-93.ArticlePubMed
- 39. Morgenstern R, Zhang J, Johansson K. Microsomal glutathione transferase 1: mechanism and functional roles. Drug Metab Rev 2011;43:300-6.ArticlePubMed
- 40. Kuang F, Liu J, Xie Y, Tang D, Kang R. MGST1 is a redox-sensitive repressor of ferroptosis in pancreatic cancer cells. Cell Chem Biol 2021;28:765-75.ArticlePubMed
- 41. Wu X, Li Y, Zhang S, Zhou X. Ferroptosis as a novel therapeutic target for cardiovascular disease. Theranostics 2021;11:3052-9.ArticlePubMedPMC
- 42. Hartmann S, Ridley AJ, Lutz S. The function of Rho-associated kinases ROCK1 and ROCK2 in the pathogenesis of cardiovascular disease. Front Pharmacol 2015;6:276.ArticlePubMedPMC
- 43. Zhang YM, Bo J, Taffet GE, Chang J, Shi J, Reddy AK, et al. Targeted deletion of ROCK1 protects the heart against pressure overload by inhibiting reactive fibrosis. FASEB J 2006;20:916-25.ArticlePubMedPDF
- 44. Rikitake Y, Oyama N, Wang CY, Noma K, Satoh M, Kim HH, et al. Decreased perivascular fibrosis but not cardiac hypertrophy in ROCK1+/- haploinsufficient mice. Circulation 2005;112:2959-65.ArticlePubMedPMC
- 45. Shimizu T, Liao JK. Rho kinases and cardiac remodeling. Circ J 2016;80:1491-8.ArticlePubMedPMC
- 46. Guan P, Liang Y, Wang N. Fasudil alleviates pressure overload-induced heart failure by activating Nrf2-mediated antioxidant responses. J Cell Biochem 2018;119:6452-60.ArticlePubMedPDF
- 47. Dong LY, Qiu XX, Zhuang Y, Xue S. Y-27632, a Rho-kinase inhibitor, attenuates myocardial ischemia-reperfusion injury in rats. Int J Mol Med 2019;43:1911-9.ArticlePubMed
- 48. Fraga-Silva RA, Sorg BS, Wankhede M, Dedeugd C, Jun JY, Baker MB, et al. ACE2 activation promotes antithrombotic activity. Mol Med 2010;16:210-5.ArticlePubMedPMCPDF
 PubReader
PubReader ePub Link
ePub Link Cite
Cite